

Lst1p and Sec24p cooperate in sorting of the plasma membrane ATPase into COPII vesicles in Saccharomyces cerevisiae
- PMID: 11086000
- PMCID: PMC2174359
- DOI: 10.1083/jcb.151.5.973
Lst1p and Sec24p cooperate in sorting of the plasma membrane ATPase into COPII vesicles in Saccharomyces cerevisiae
Abstract
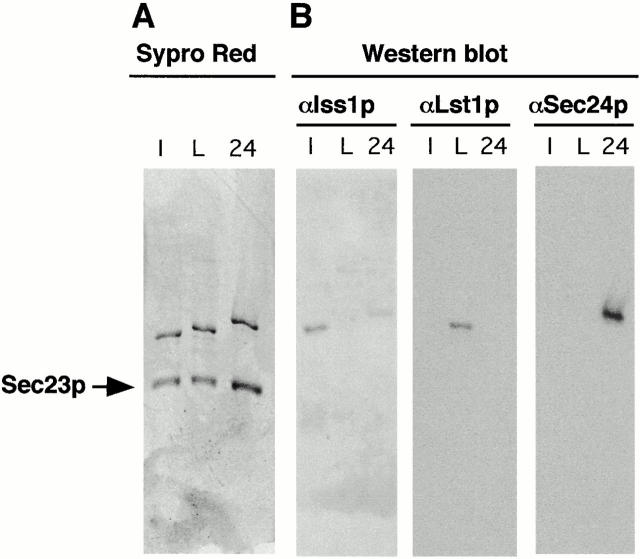
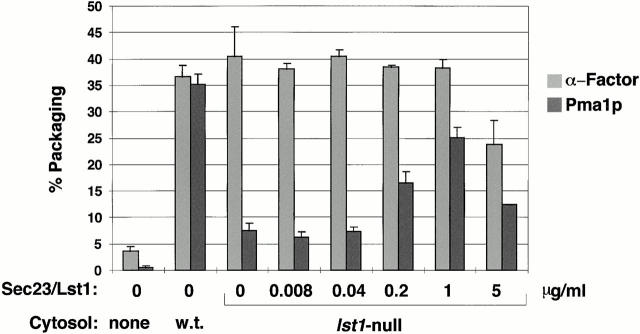
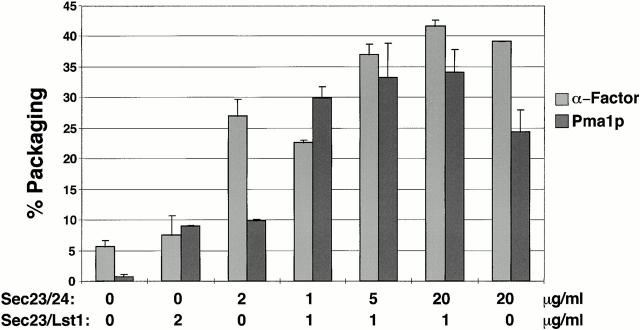
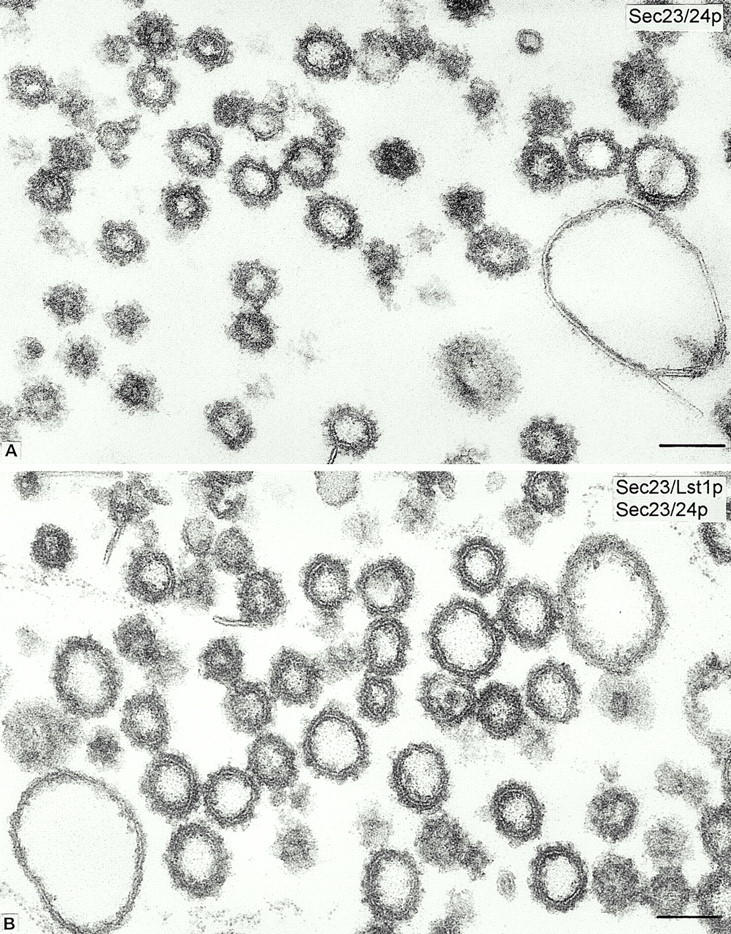
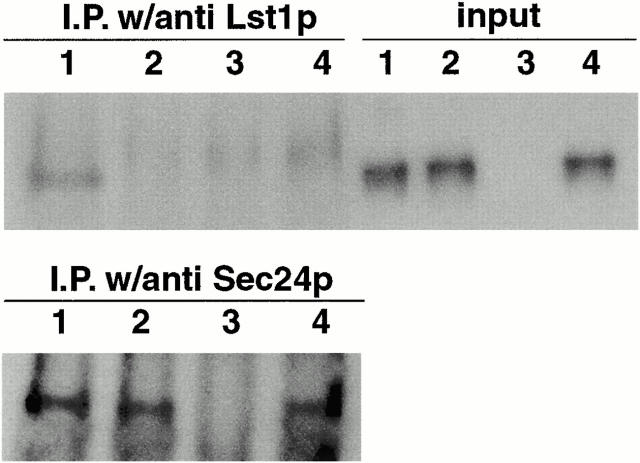
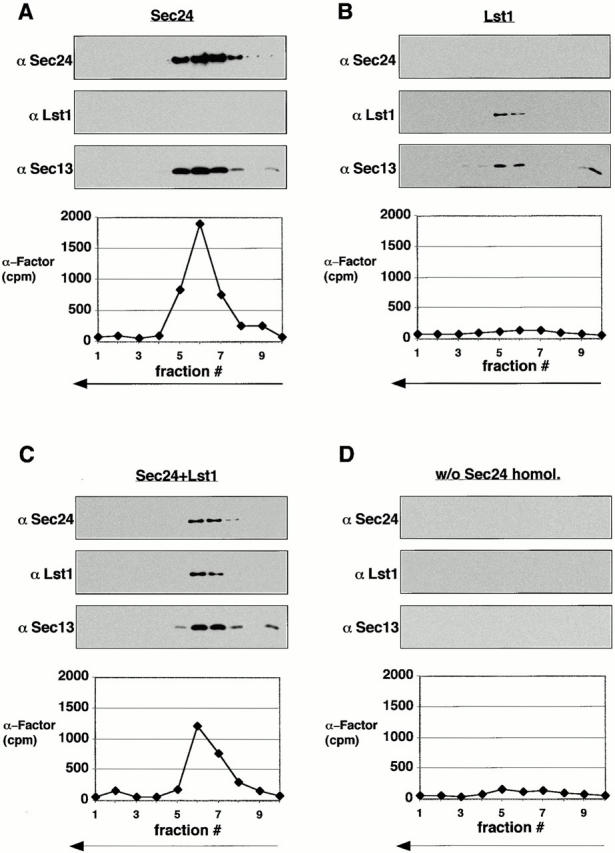
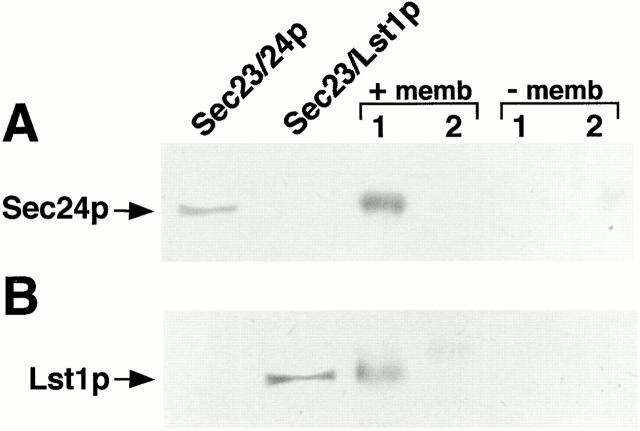
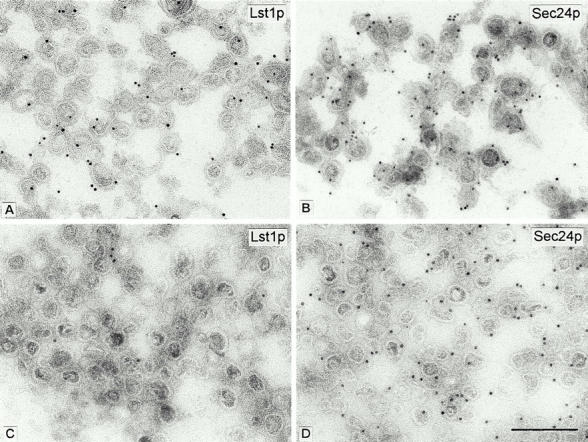
Formation of ER-derived protein transport vesicles requires three cytosolic components, a small GTPase, Sar1p, and two heterodimeric complexes, Sec23/24p and Sec13/31p, which comprise the COPII coat. We investigated the role of Lst1p, a Sec24p homologue, in cargo recruitment into COPII vesicles in Saccharomyces cerevisiae. A tagged version of Lst1p was purified and eluted as a heterodimer complexed with Sec23p comparable to the Sec23/24p heterodimer. We found that cytosol from an lst1-null strain supported the packaging of alpha-factor precursor into COPII vesicles but was deficient in the packaging of Pma1p, the essential plasma membrane ATPase. Supplementation of mutant cytosol with purified Sec23/Lst1p restored Pma1p packaging into the vesicles. When purified COPII components were used in the vesicle budding reaction, Pma1p packaging was optimal with a mixture of Sec23/24p and Sec23/Lst1p; Sec23/Lst1p did not replace Sec23/24p. Furthermore, Pma1p coimmunoprecipitated with Lst1p and Sec24p from vesicles. Vesicles formed with a mixture of Sec23/Lst1p and Sec23/24p were similar morphologically and in their buoyant density, but larger than normal COPII vesicles (87-nm vs. 75-nm diameter). Immunoelectronmicroscopic and biochemical studies revealed both Sec23/Lst1p and Sec23/24p on the membranes of the same vesicles. These results suggest that Lst1p and Sec24p cooperate in the packaging of Pma1p and support the view that biosynthetic precursors of plasma membrane proteins must be sorted into ER-derived transport vesicles. Sec24p homologues may comprise a more complex coat whose combinatorial subunit composition serves to expand the range of cargo to be packaged into COPII vesicles. By changing the geometry of COPII coat polymerization, Lst1p may allow the transport of bulky cargo molecules, polymers, or particles.
Figures
References
-
- Auer M., Scarborough G.A., Kühlbrandt W. Three-dimensional map of the plasma membrane H+-ATPase in the open conformation. Nature. 1998;392:840–843. - PubMed
-
- Baker D., Hicke L., Rexach M., Schleyer M., Schekman R. Reconstitution of SEC gene product-dependent intercompartmental protein transport. Cell. 1988;54:335–344. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
