

Region-specific microtubule transport in motile cells
- PMID: 11086002
- PMCID: PMC2174357
- DOI: 10.1083/jcb.151.5.1003
Region-specific microtubule transport in motile cells
Abstract
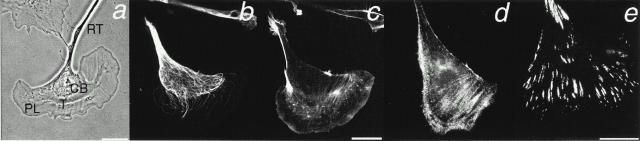
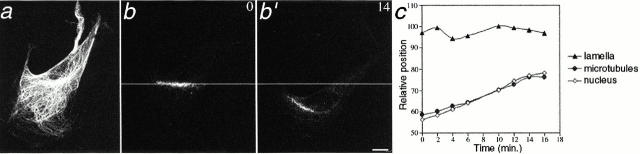
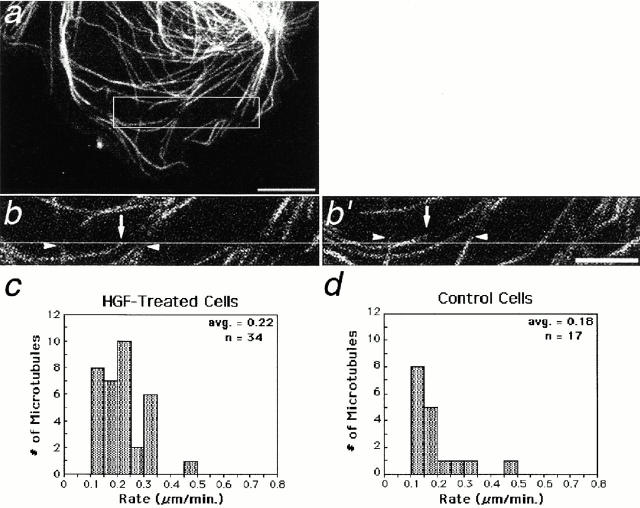
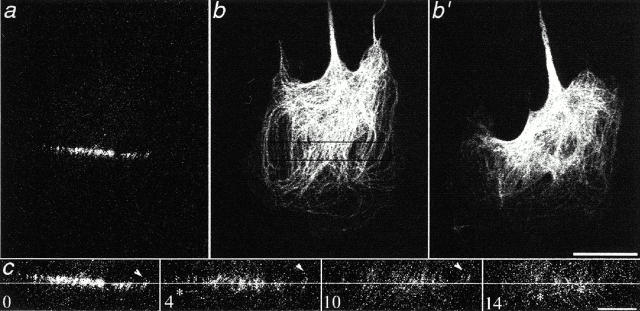
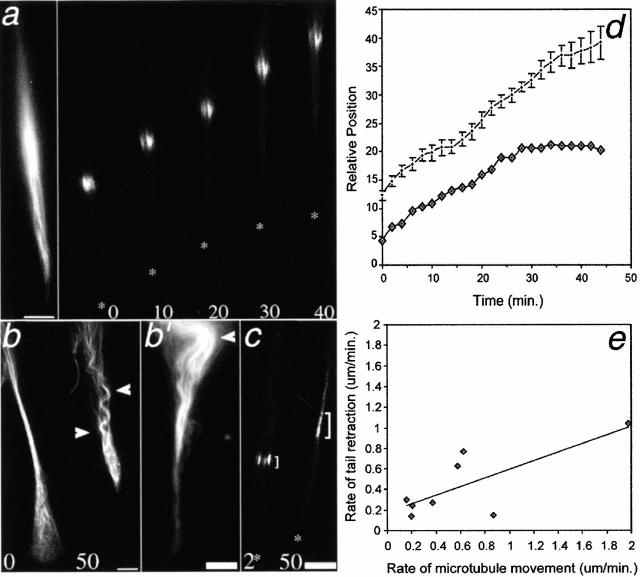
Photoactivation and photobleaching of fluorescence were used to determine the mechanism by which microtubules (MTs) are remodeled in PtK2 cells during fibroblast-like motility in response to hepatocyte growth factor (HGF). The data show that MTs are transported during cell motility in an actomyosin-dependent manner, and that the direction of transport depends on the dominant force in the region examined. MTs in the leading lamella move rearward relative to the substrate, as has been reported in newt cells (Waterman-Storer, C.M., and E.D. Salmon. 1997. J. Cell Biol. 139:417-434), whereas MTs in the cell body and in the retraction tail move forward, in the direction of cell locomotion. In the transition zone between the peripheral lamella and the cell body, a subset of MTs remains stationary with respect to the substrate, whereas neighboring MTs are transported either forward, with the cell body, or rearward, with actomyosin retrograde flow. In addition to transport, the photoactivated region frequently broadens, indicating that individual marked MTs are moved either at different rates or in different directions. Mark broadening is also observed in nonmotile cells, indicating that this aspect of transport is independent of cell locomotion. Quantitative measurements of the dissipation of photoactivated fluorescence show that, compared with MTs in control nonmotile cells, MT turnover is increased twofold in the lamella of HGF-treated cells but unchanged in the retraction tail, demonstrating that microtubule turnover is regionally regulated.
Figures

References
-
- Baas P.W., Brown A. Slow axonal transportthe polymer transport model. Trends Cell Biol. 1997;7:380–384. - PubMed
-
- Danowski B. Fibroblast contractility and actin organization are stimulated by microtubule inhibitors. J. Cell Sci. 1989;93:255–266. - PubMed
-
- Desai A., Mitchinson T.J. Preparation and characterization of caged fluorescein tubulin. Methods Enzymol. 1998;298:128–1320. - PubMed
-
- Euteneuer U., Schliwa M. Persistent, directional motility of cells and cytoplasmic fragments in the absence of microtubules. Nature. 1984;310:58–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
