

A role for ubiquitin ligase recruitment in retrovirus release
- PMID: 11087860
- PMCID: PMC27178
- DOI: 10.1073/pnas.97.24.13063
A role for ubiquitin ligase recruitment in retrovirus release
Abstract
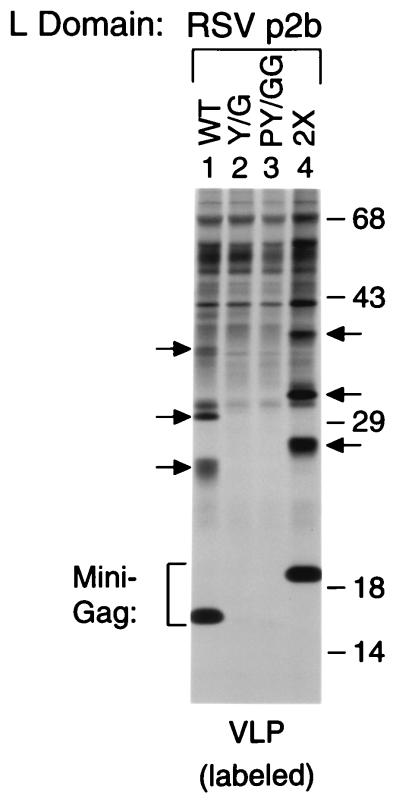
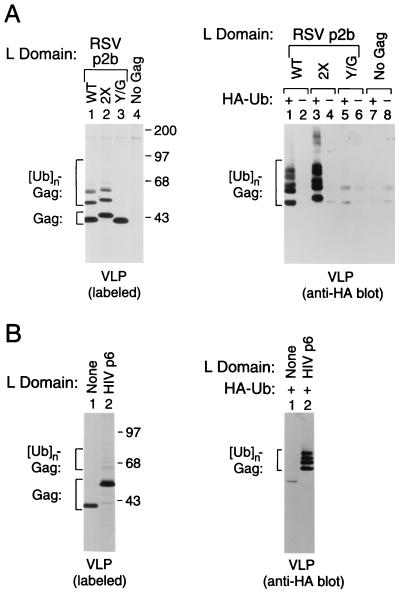
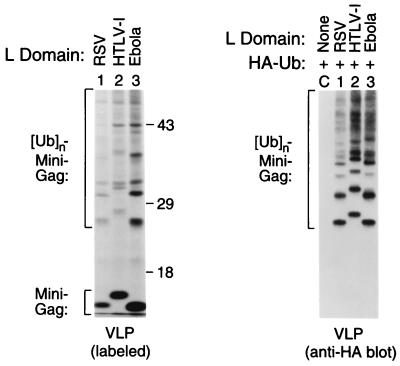
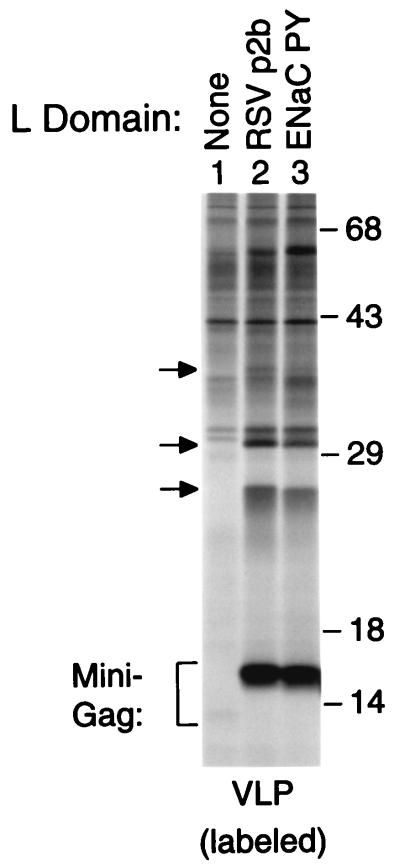
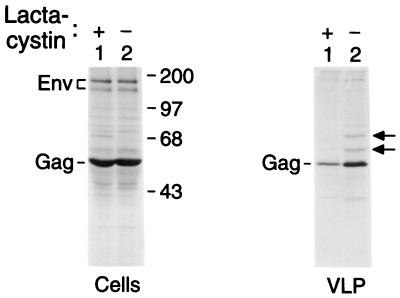
Retroviral Gag polyproteins have specific regions, commonly referred to as late assembly (L) domains, which are required for the efficient separation of assembled virions from the host cell. The L domain of HIV-1 is in the C-terminal p6(gag) domain and contains an essential P(T/S)AP core motif that is widely conserved among lentiviruses. In contrast, the L domains of oncoretroviruses such as Rous sarcoma virus (RSV) have a more N-terminal location and a PPxY core motif. In the present study, we used chimeric Gag constructs to probe for L domain activity, and observed that the unrelated L domains of RSV and HIV-1 both induced the appearance of Gag-ubiquitin conjugates in virus-like particles (VLP). Furthermore, a single-amino acid substitution that abolished the activity of the RSV L domain in VLP release also abrogated its ability to induce Gag ubiquitination. Particularly robust Gag ubiquitination and enhancement of VLP release were observed in the presence of the candidate L domain of Ebola virus, which contains overlapping P(T/S)AP and PPxY motifs. The release defect of a minimal Gag construct could also be corrected through the attachment of a peptide that serves as a physiological docking site for the ubiquitin ligase Nedd4. Furthermore, VLP formation by a full-length Gag polyprotein was sensitive to lactacystin, which depletes the levels of free ubiquitin through inhibition of the proteasome. Our findings suggest that the engagement of the ubiquitin conjugation machinery by L domains plays a crucial role in the release of a diverse group of enveloped viruses.
Figures
Comment in
-
Ubiquitin in retrovirus assembly: actor or bystander?Proc Natl Acad Sci U S A. 2000 Nov 21;97(24):12945-7. doi: 10.1073/pnas.97.24.12945. Proc Natl Acad Sci U S A. 2000. PMID: 11087848 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
