

A null mutation in the UL36 gene of herpes simplex virus type 1 results in accumulation of unenveloped DNA-filled capsids in the cytoplasm of infected cells
- PMID: 11090159
- PMCID: PMC112442
- DOI: 10.1128/jvi.74.24.11608-11618.2000
A null mutation in the UL36 gene of herpes simplex virus type 1 results in accumulation of unenveloped DNA-filled capsids in the cytoplasm of infected cells
Abstract
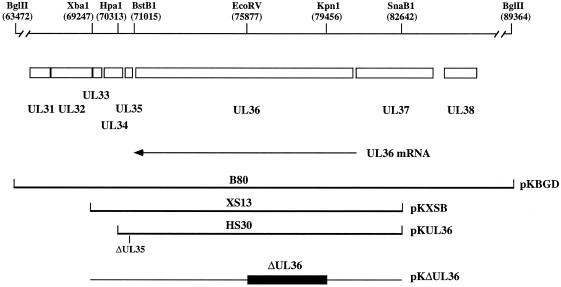
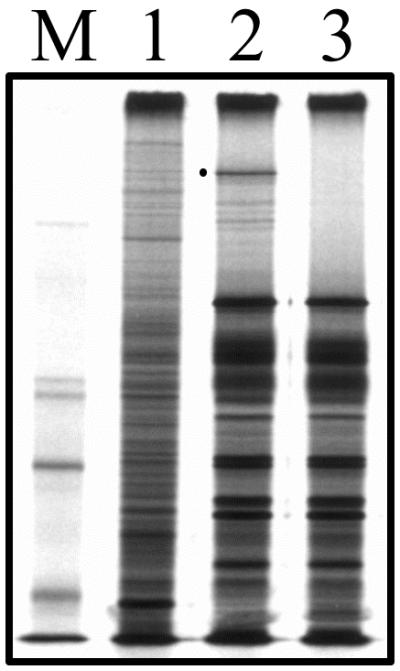
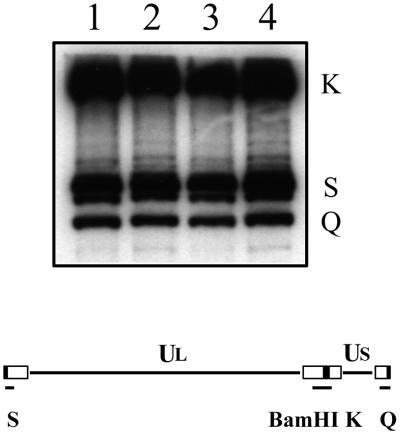
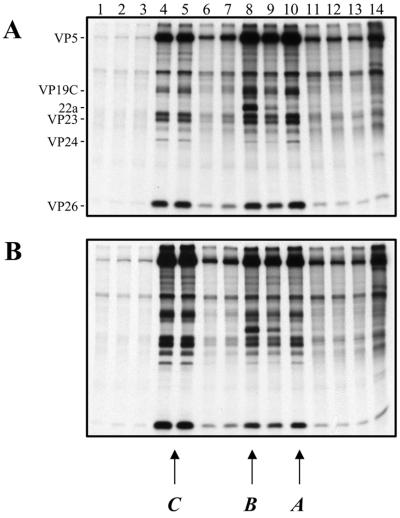
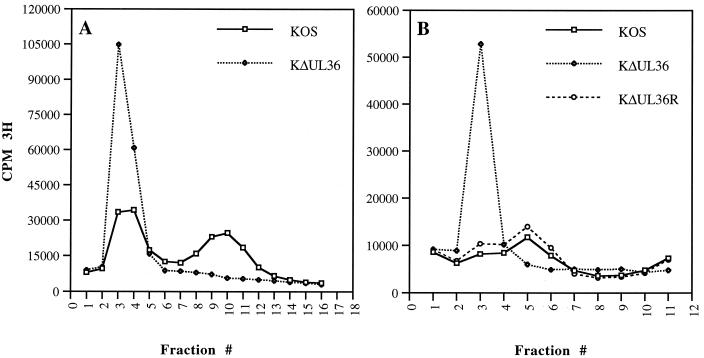
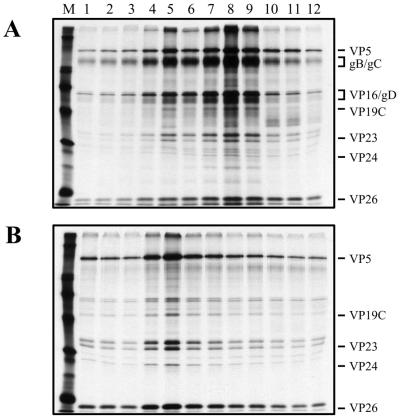
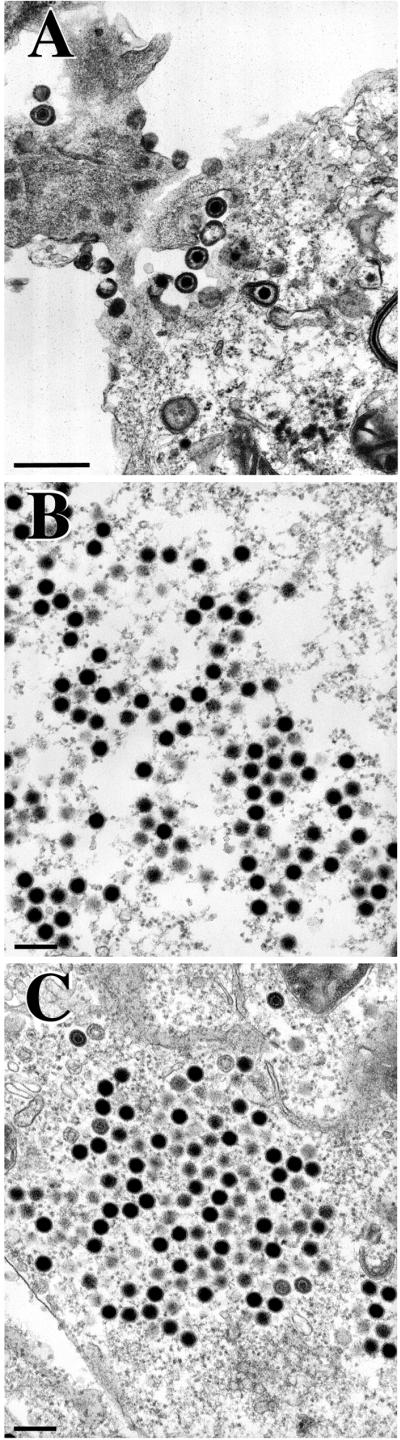
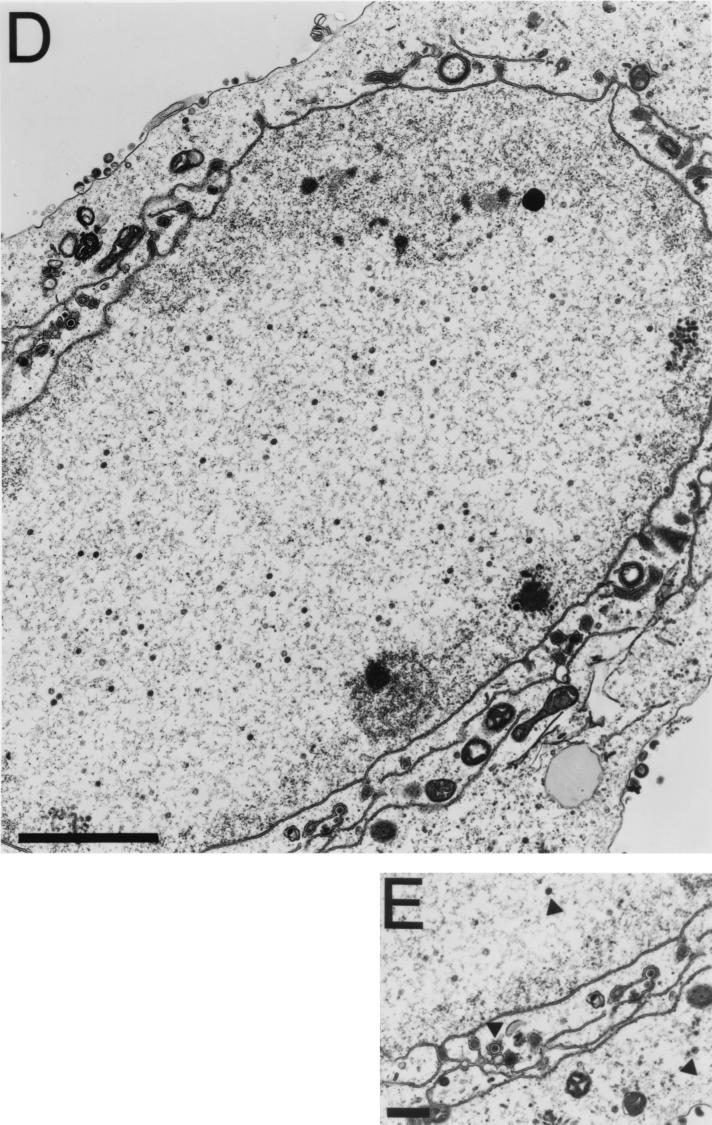
The UL36 open reading frame (ORF) encodes the largest herpes simplex virus type 1 (HSV-1) protein, a 270-kDa polypeptide designated VP1/2, which is also a component of the virion tegument. A null mutation was generated in the UL36 gene to elucidate its role in the virus life cycle. Since the UL36 gene specifies an essential function, complementing cell lines transformed for sequences encoding the UL36 ORF were made. A mutant virus, designated KDeltaUL36, that encodes a null mutation in the UL36 gene was isolated and propagated in these cell lines. When noncomplementing cells infected with KDeltaUL36 were analyzed, both terminal genomic DNA fragments and DNA-containing capsids (C capsids) were detected; therefore, UL36 is not required for cleavage or packaging of DNA. Sedimentation analysis of lysates from mutant-infected cells revealed the presence of particles that have the physical characteristics of C capsids. In agreement with this, polypeptide profiles of the mutant particles revealed an absence of the major envelope and tegument components. Ultrastructural analysis revealed the presence of numerous unenveloped DNA containing capsids in the cytoplasm of KDeltaUL36-infected cells. The UL36 mutant particles were tagged with the VP26-green fluorescent protein marker, and their movement was monitored in living cells. In KDeltaUL36-infected cells, extensive particulate fluorescence corresponding to the capsid particles was observed throughout the cytosol. Accumulation of fluorescence at the plasma membrane which indicated maturation and egress of virions was observed in wild-type-infected cells but was absent in KDeltaUL36-infected cells. In the absence of UL36 function, DNA-filled capsids are produced; these capsids enter the cytosol after traversing the nuclear envelope and do not mature into enveloped virus. The maturation and egress of the UL36 mutant particles are abrogated, possibly due to a late function of this complex polypeptide, i.e., to target capsids to the correct maturation pathway.
Figures
References
-
- Ace C I, Dalrymple M A, Ramsay F H, Preston V G, Preston C M. Mutational analysis of the herpes simplex virus type 1 trans-inducing factor Vmw65. J Gen Virol. 1988;69:2595–2605. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
