

Lack of shielding of primer binding site silencer-mediated repression of an internal promoter in a retrovirus vector by the putative insulators scs, BEAD-1, and HS4
- PMID: 11090169
- PMCID: PMC112452
- DOI: 10.1128/jvi.74.24.11697-11707.2000
Lack of shielding of primer binding site silencer-mediated repression of an internal promoter in a retrovirus vector by the putative insulators scs, BEAD-1, and HS4
Abstract
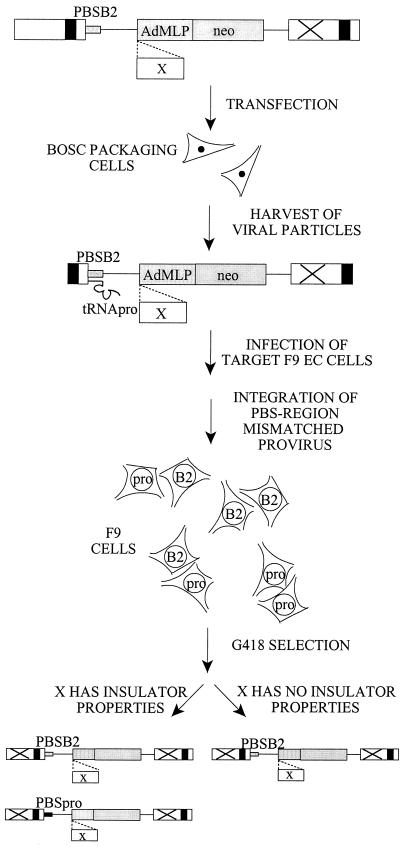
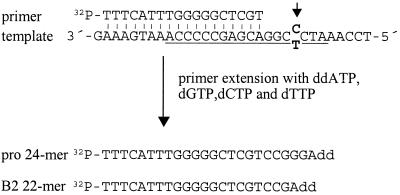
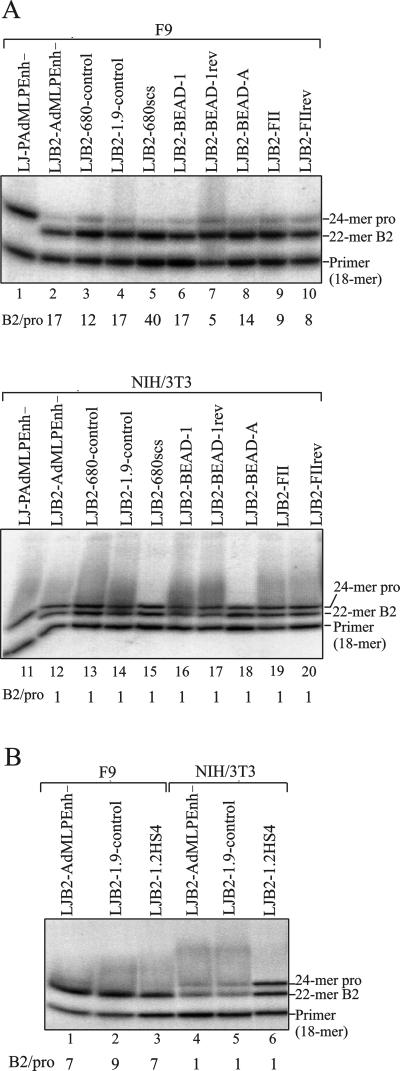
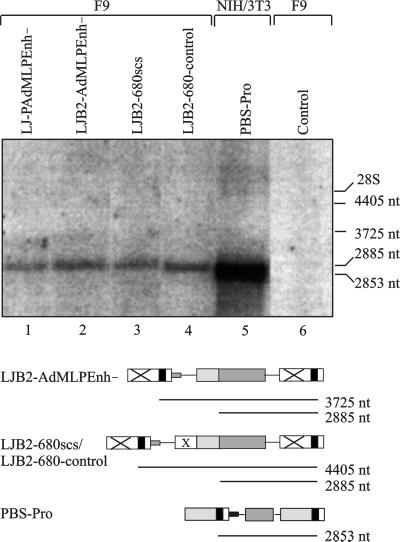
A major determinant for transcriptional incompetence of murine leukemia virus (MLV) and MLV-derived vectors in embryonal cells is located at the proline primer binding site (PBS). The mechanism of silencing is unknown, yet the effect is capable of spreading to adjacent promoters. Based on a retroviral vector containing an internal promoter and the escape mutant B2 PBS with expressional capacity in embryonal cells, we have developed an assay to test the ability of putative insulators to shield the silencer at the PBS. Since the B2 PBS reverts to the wild-type PBS at high frequency, a shielding ability of a putative insulator can be assessed from the ratio of expressing B2 PBS to proline PBS proviruses in the target embryonal carcinoma cell population as measured by primer extension. Our results show that none of the possible insulators, scs, BEAD-1, or HS4, is able to shield an internal promoter from the repressive effect of the silencer at the PBS region when inserted between the silencer and the promoter.
Figures

References
-
- Baniahmad A, Steiner C, Köhne A C, Renkawitz R. Modular structure of a chicken lysozyme silencer: involvement of an unusual thyroid hormone receptor binding site. Cell. 1990;61:505–514. - PubMed
-
- Barklis E, Mulligan R C, Jaenisch R. Chromosomal position or virus mutation permits retrovirus expression in embryonal carcinoma cells. Cell. 1986;47:391–399. - PubMed
-
- Beck E, Ludwig G, Auerswald E A, Reiss B, Schaller H. Nucleotide sequence and exact localization of the neomycin phosphotransferase gene from transposon Tn5. Gene. 1982;19:327–336. - PubMed
-
- Bell A C, West A D, Felsenfeld G. The protein CTCF is required for the enhancer blocking activity of vertebrate insulators. Cell. 1999;98:387–396. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
