

Mechanism of interference mediated by human parainfluenza virus type 3 infection
- PMID: 11090179
- PMCID: PMC112462
- DOI: 10.1128/jvi.74.24.11792-11799.2000
Mechanism of interference mediated by human parainfluenza virus type 3 infection
Abstract
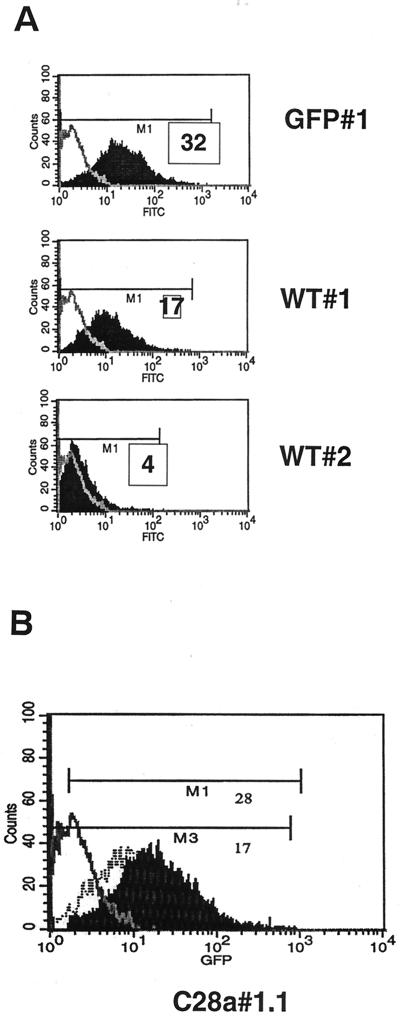
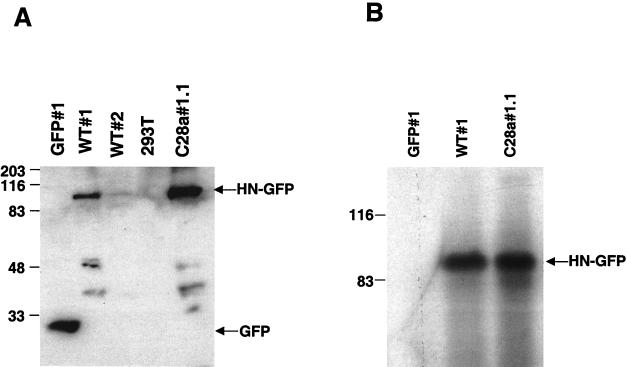
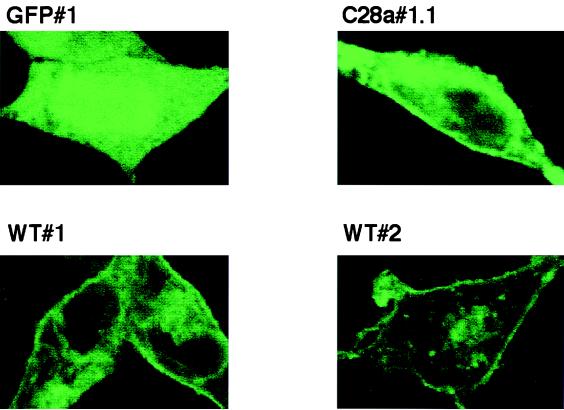
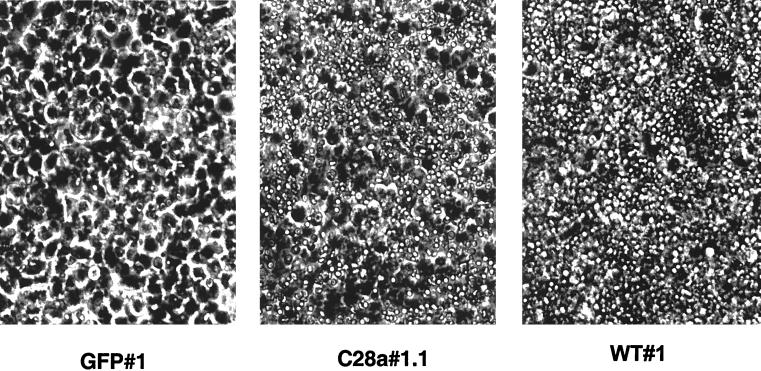
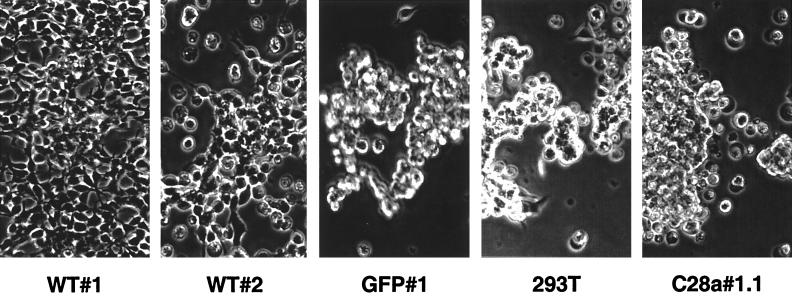
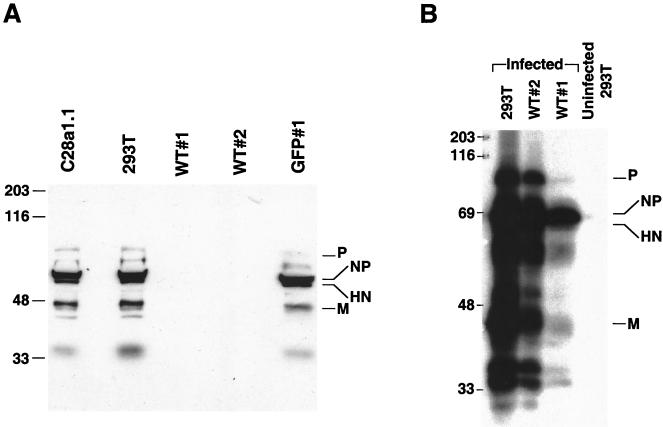
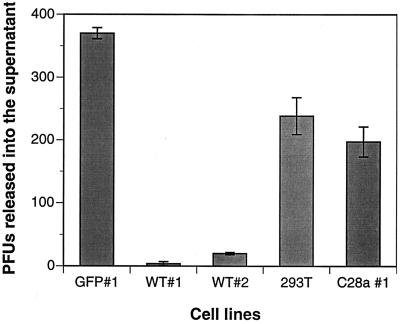
Viral interference is characterized by the resistance of infected cells to infection by a challenge virus. Mechanisms of viral interference have not been characterized for human parainfluenza virus type 3 (HPF3), and the possible role of the neuraminidase (receptor-destroying) enzyme of the hemagglutinin-neuraminidase (HN) glycoprotein has not been assessed. To determine whether continual HN expression results in depletion of the viral receptors and thus prevents entry and cell fusion, we tested whether cells expressing wild-type HPF3 HN are resistant to viral infection. Stable expression of wild-type HN-green fluorescent protein (GFP) on cell membranes in different amounts allowed us to establish a correlation between the level of HN expression, the level of neuraminidase activity, and the level of protection from HPF3 infection. Cells with the highest levels of HN expression and neuraminidase activity on the cell surface were most resistant to infection by HPF3. To determine whether this resistance is attributable to the viral neuraminidase, we used a cloned variant HPF3 HN that has two amino acid alterations in HN leading to the loss of detectable neuraminidase activity. Cells expressing the neuraminidase-deficient variant HN-GFP were not protected from infection, despite expressing HN on their surface at levels even higher than the wild-type cell clones. Our results demonstrate that the HPF3 HN-mediated interference effect can be attributed to the presence of an active neuraminidase enzyme activity and provide the first definitive evidence that the mechanism for attachment interference by a paramyxovirus is attributable to the viral neuraminidase.
Figures
References
-
- Ah-Tye C, Schwartz S, Huberman K, Carlin E, Moscona A. Virus-receptor interactions of human parainfluenza viruses types 1, 2 and 3. Microb Pathog. 1999;27:329–336. - PubMed
-
- Baluda M A. Homologous interference by ultraviolet-inactivated Newcastle disease virus. Virology. 1957;7:315–327. - PubMed
-
- Bratt M A, Rubin H. Specific interference among strains of NDV. I. Demonstration and measurement of interference. Virology. 1967;33:598–608. - PubMed
-
- Bratt M A, Rubin H. Specific interference among strains of NDV. II. Comparison of interference of active and inactive virus. Virology. 1967;35:381–394. - PubMed
-
- Bratt M A, Rubin H. Specific interference among strains of NDV. III. Mechanism of interference. Virology. 1968;35:395–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
