

Oriented asymmetric divisions that generate the stomatal spacing pattern in arabidopsis are disrupted by the too many mouths mutation
- PMID: 11090210
- PMCID: PMC150159
- DOI: 10.1105/tpc.12.11.2075
Oriented asymmetric divisions that generate the stomatal spacing pattern in arabidopsis are disrupted by the too many mouths mutation
Abstract
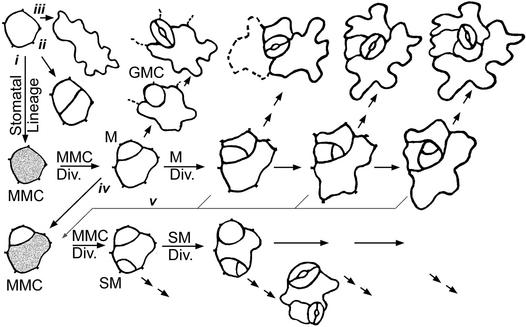
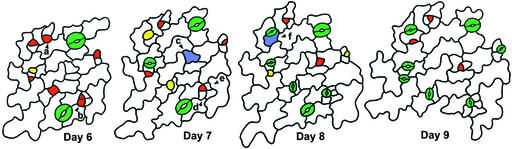
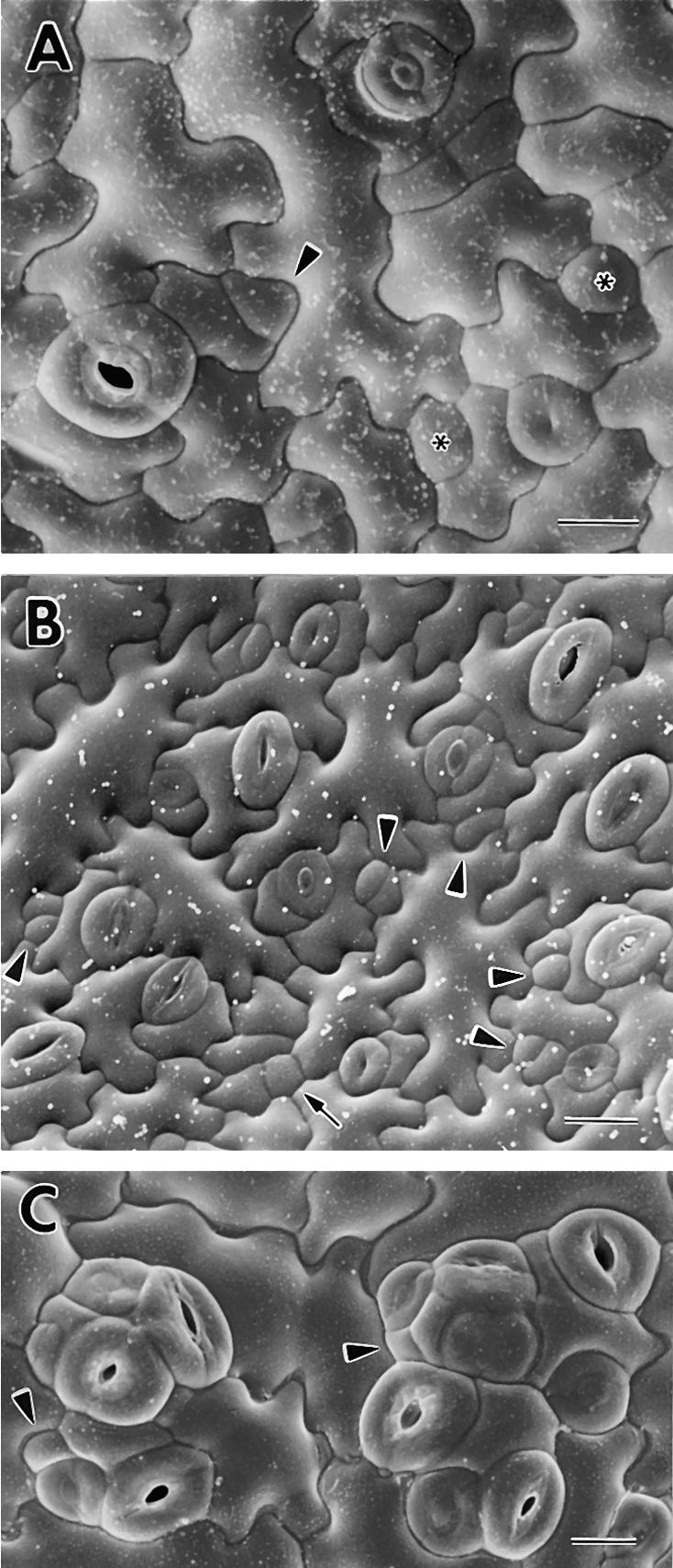
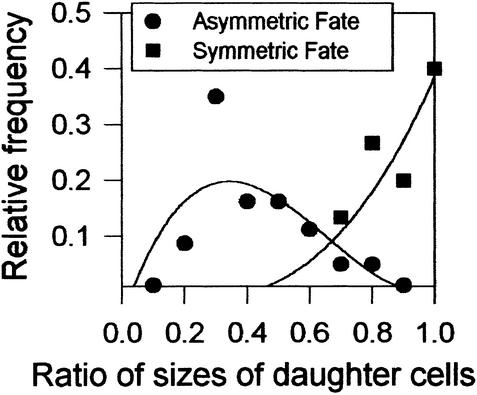
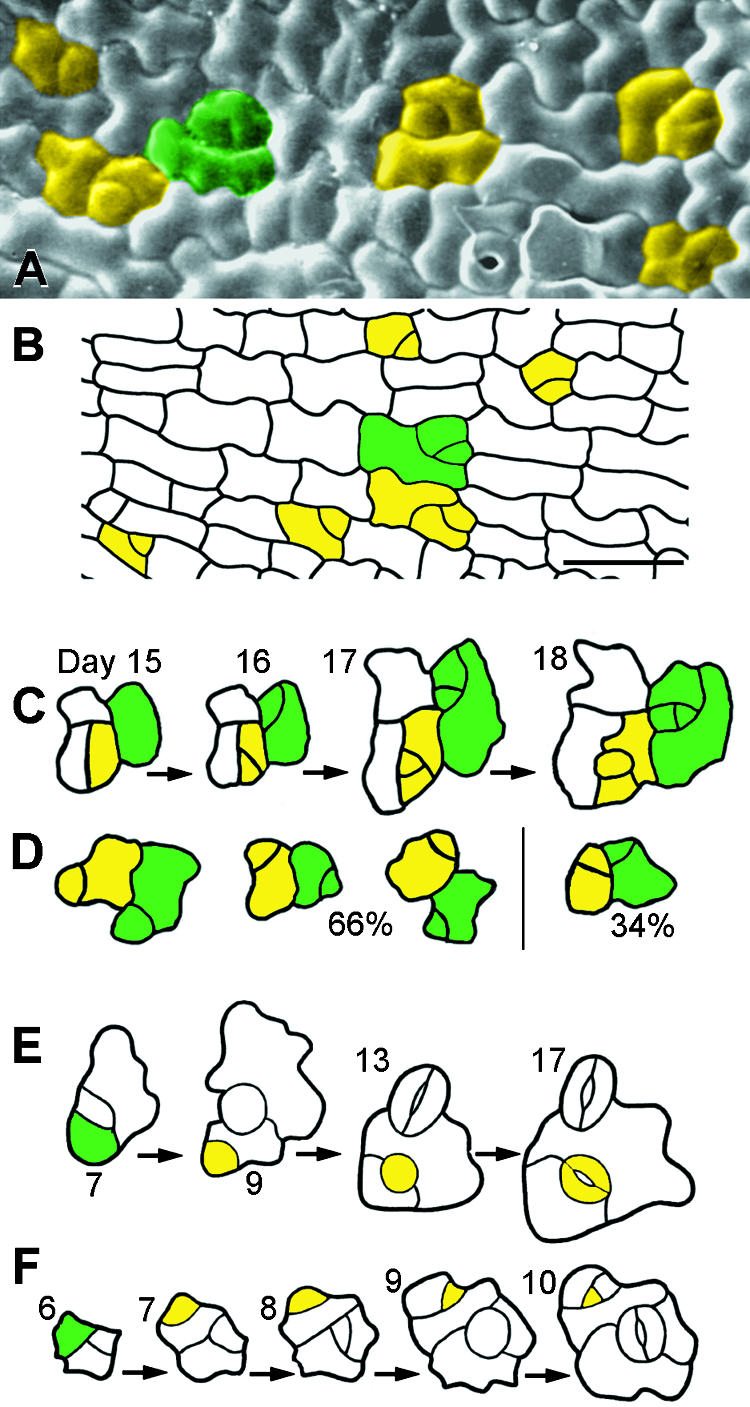
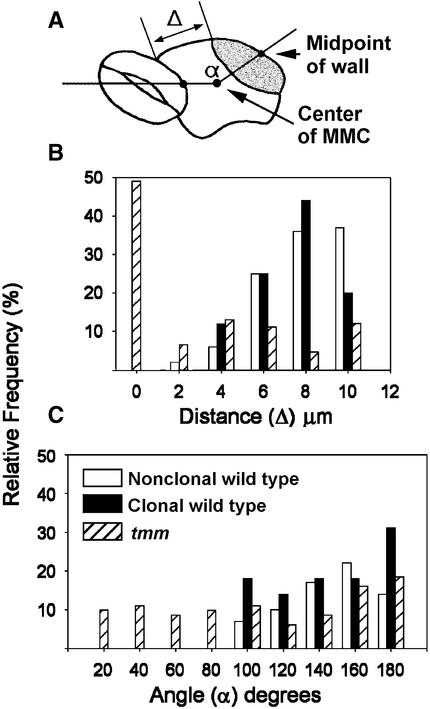
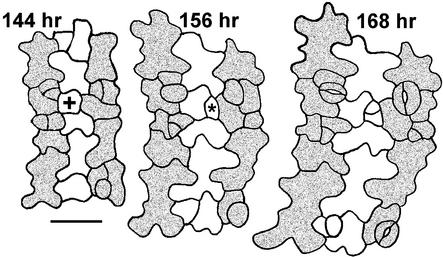
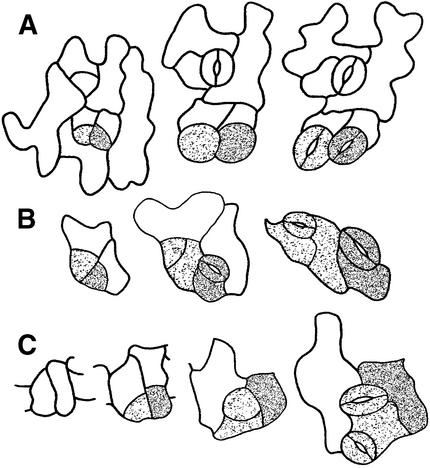
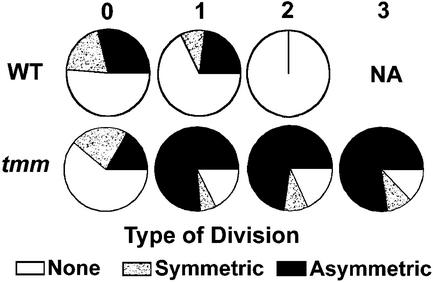
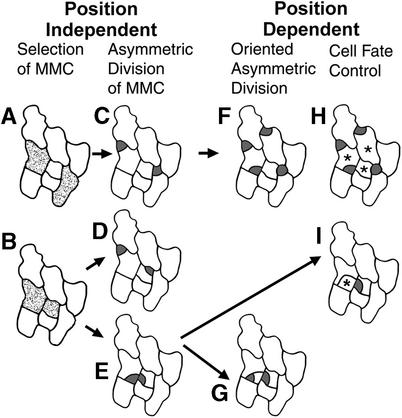
Wild-type stomata are spaced by intervening cells, a pattern disrupted in the Arabidopsis mutant too many mouths (tmm). To determine the mechanism of wild-type spacing and how tmm results in pattern violations, we analyzed the behavior of cells through time by using sequential dental resin impressions. Meristemoids are stomatal precursors produced by asymmetric division. We show that wild-type patterning largely results when divisions next to a preexisting stoma or precursor are oriented so that the new meristemoid is placed away. Because this placement is independent of cell lineage, these divisions may be oriented by cell-cell signaling. tmm randomizes this orientation and releases a prohibition on asymmetric division in cells at specific locations, resulting in stomatal clusters. TMM is thus necessary for two position-dependent events in leaves: the orientation of asymmetric divisions that pattern stomata, and the control of which cells will enter the stomatal pathway. In addition, our findings argue against most previous hypotheses of wild-type stomatal patterning.
Figures

References
-
- Bünning, E. (1956). General processes of differentiation. In The Growth of Leaves, F.L. Milthorpe, ed (London: Butterworths Scientific Publications), pp. 18–30.
-
- Croxdale, J.L. (2000). Stomatal patterning in angiosperms. Am. J. Bot. 87, 1069–1080. - PubMed
-
- Drubin, D.G., and Nelson, W.J.. (1996). Origins of cell polarity. Cell 84, 335–344. - PubMed
-
- Fowler, J.E., and Quatrano, R.S. (1997). Plant cell morphogenesis: Plasma membrane interactions with the cytoskeleton and cell wall. Annu. Rev. Cell Dev. Biol. 13, 697–743. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
