

Molecular evolution of virulence in natural field strains of Xanthomonas campestris pv. vesicatoria
- PMID: 11092868
- PMCID: PMC94833
- DOI: 10.1128/JB.182.24.7053-7059.2000
Molecular evolution of virulence in natural field strains of Xanthomonas campestris pv. vesicatoria
Abstract
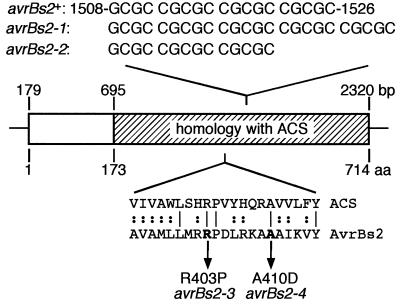
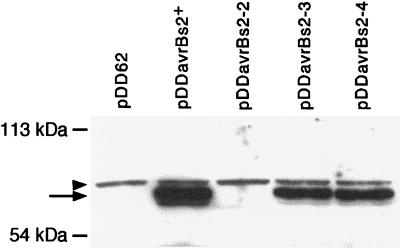
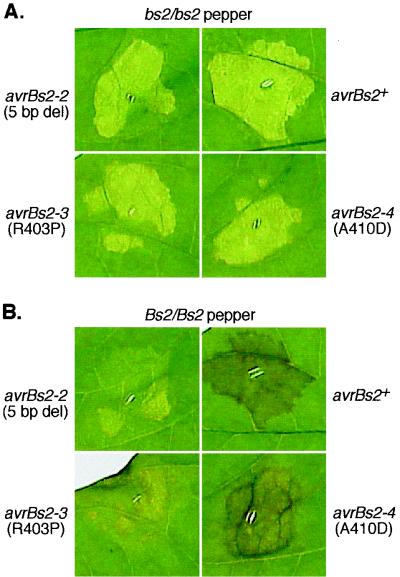
The avrBs2 avirulence gene of the bacterial plant pathogen Xanthomonas campestris pv. vesicatoria triggers disease resistance in pepper plants containing the Bs2 resistance gene and contributes to bacterial virulence on susceptible host plants. We studied the effects of the pepper Bs2 gene on the evolution of avrBs2 by characterizing the molecular basis for virulence of 20 X. campestris pv. vesicatoria field strains that were isolated from disease spots on previously resistant Bs2 pepper plants. All field strains tested were complemented by a wild-type copy of avrBs2 in their ability to trigger disease resistance on Bs2 plants. DNA sequencing revealed four mutant alleles of avrBs2, two of which consisted of insertions or deletions of 5 nucleotides in a repetitive region of avrBs2. The other two avrBs2 alleles were characterized by point mutations with resulting single amino acid changes (R403P or A410D). We generated isogenic X. campestris pv. vesicatoria strains by chromosomal avrBs2 gene exchange to study the effects of these mutations on the dual functions of avrBs2 in enhancing bacterial virulence and inducing plant resistance by in planta bacterial growth experiments. The deletion of 5 nucleotides led to loss of avrBs2-induced resistance on Bs2 pepper plants and abolition of avrBs2-mediated enhancement of fitness on susceptible plants. Significantly, the point mutations led to minimal reduction in virulence function of avrBs2 on susceptible pepper plants, with either minimal (R403P allele) or an intermediate level of (A410D allele) triggering of resistance on Bs2 plants. Consistent with the divergent selection pressures on avrBs2 exerted by the Bs2 resistance gene, our results show that avrBs2 is evolving to decrease detection by the Bs2 gene while at the same time maintaining its virulence function.
Figures
Similar articles
-
Spontaneous and induced mutations in a single open reading frame alter both virulence and avirulence in Xanthomonas campestris pv. vesicatoria avrBs2.J Bacteriol. 1996 Aug;178(15):4661-9. doi: 10.1128/jb.178.15.4661-4669.1996. J Bacteriol. 1996. PMID: 8755898 Free PMC article.
-
Inducible expression of Bs2 R gene from Capsicum chacoense in sweet orange (Citrus sinensis L. Osbeck) confers enhanced resistance to citrus canker disease.Plant Mol Biol. 2017 Apr;93(6):607-621. doi: 10.1007/s11103-017-0586-8. Epub 2017 Feb 2. Plant Mol Biol. 2017. PMID: 28155188
-
Molecular signals required for type III secretion and translocation of the Xanthomonas campestris AvrBs2 protein to pepper plants.Proc Natl Acad Sci U S A. 2000 Nov 21;97(24):13324-9. doi: 10.1073/pnas.230450797. Proc Natl Acad Sci U S A. 2000. PMID: 11078519 Free PMC article.
-
Genomic approaches in Xanthomonas campestris pv. vesicatoria allow fishing for virulence genes.J Biotechnol. 2003 Dec 19;106(2-3):203-14. doi: 10.1016/j.jbiotec.2003.07.012. J Biotechnol. 2003. PMID: 14651862 Review.
-
Molecular functions of Xanthomonas type III effector AvrBsT and its plant interactors in cell death and defense signaling.Planta. 2017 Feb;245(2):237-253. doi: 10.1007/s00425-016-2628-x. Epub 2016 Dec 7. Planta. 2017. PMID: 27928637 Review.
Cited by
-
The role of type III effectors from Xanthomonas axonopodis pv. manihotis in virulence and suppression of plant immunity.Mol Plant Pathol. 2018 Mar;19(3):593-606. doi: 10.1111/mpp.12545. Epub 2017 Apr 5. Mol Plant Pathol. 2018. PMID: 28218447 Free PMC article.
-
The type III effectors of Xanthomonas.Mol Plant Pathol. 2009 Nov;10(6):749-66. doi: 10.1111/j.1364-3703.2009.00590.x. Mol Plant Pathol. 2009. PMID: 19849782 Free PMC article. Review.
-
Xanthomonas T3S Effector XopN Suppresses PAMP-Triggered Immunity and Interacts with a Tomato Atypical Receptor-Like Kinase and TFT1.Plant Cell. 2009 Apr;21(4):1305-23. doi: 10.1105/tpc.108.063123. Epub 2009 Apr 14. Plant Cell. 2009. PMID: 19366901 Free PMC article.
-
The N-terminal region of Pseudomonas type III effector AvrPtoB elicits Pto-dependent immunity and has two distinct virulence determinants.Plant J. 2007 Nov;52(4):595-614. doi: 10.1111/j.1365-313X.2007.03259.x. Epub 2007 Aug 31. Plant J. 2007. PMID: 17764515 Free PMC article.
-
Transgenic Improvement for Biotic Resistance of Crops.Int J Mol Sci. 2022 Nov 19;23(22):14370. doi: 10.3390/ijms232214370. Int J Mol Sci. 2022. PMID: 36430848 Free PMC article. Review.
References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J G, Struhl K. Current protocols in molecular biology. New York, N.Y: Greene Publishing Associates and Wiley-Interscience; 1987.
-
- Dangl J L. The enigmatic avirulence genes of phytopathogenic bacteria. In: Dangl J L, editor. Bacterial pathogenesis of plants and animals—molecular and cellular mechanisms. Berlin, Germany: Springer-Verlag; 1994. pp. 99–118. - PubMed
-
- Daniels M J, Barber C E, Turner D C, Cleary W G, Sawczyc M K. Isolation of mutants of Xanthomonas campestris pv. campestris with altered pathogenicity. J Gen Microbiol. 1984;130:2447–2455.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
