

The MN1-TEL fusion protein, encoded by the translocation (12;22)(p13;q11) in myeloid leukemia, is a transcription factor with transforming activity
- PMID: 11094079
- PMCID: PMC102185
- DOI: 10.1128/MCB.20.24.9281-9293.2000
The MN1-TEL fusion protein, encoded by the translocation (12;22)(p13;q11) in myeloid leukemia, is a transcription factor with transforming activity
Abstract
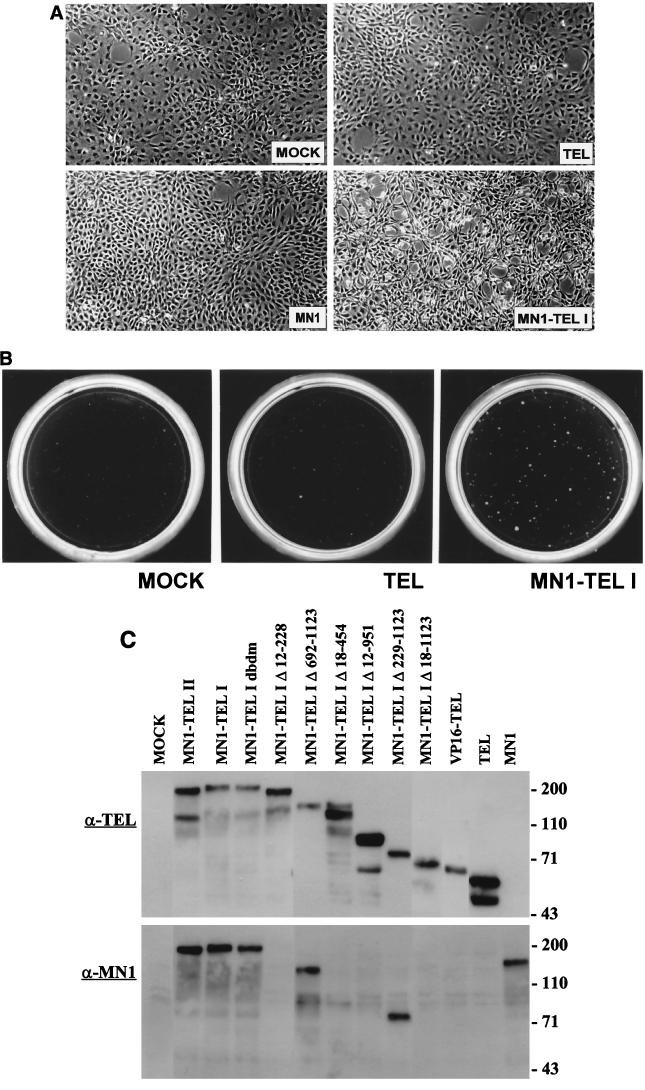
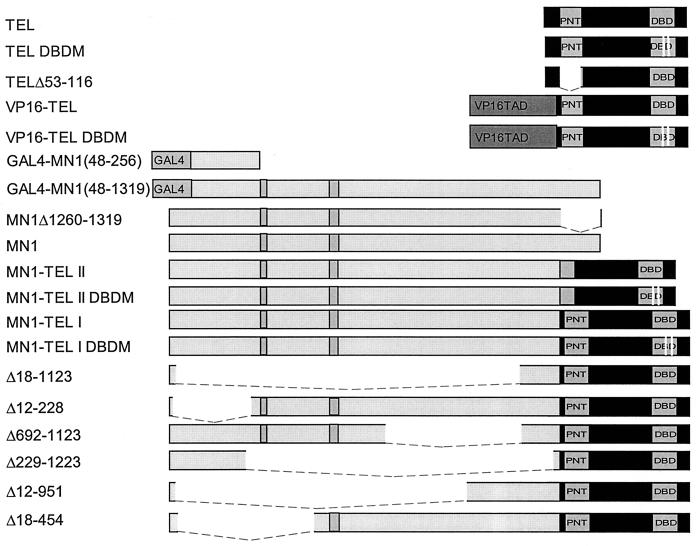
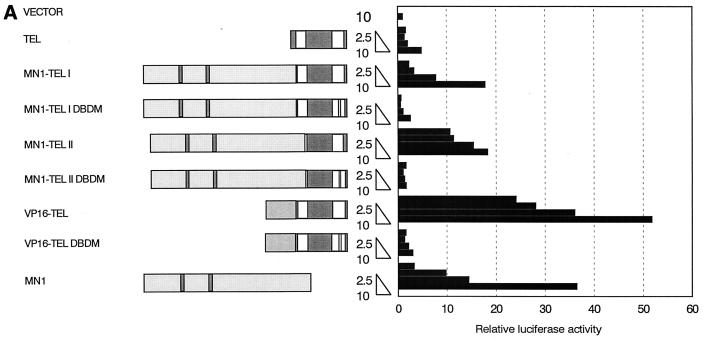
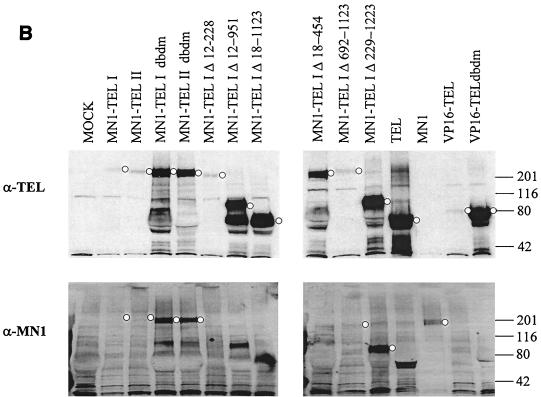
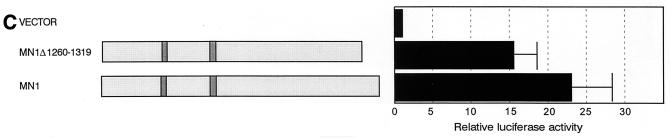
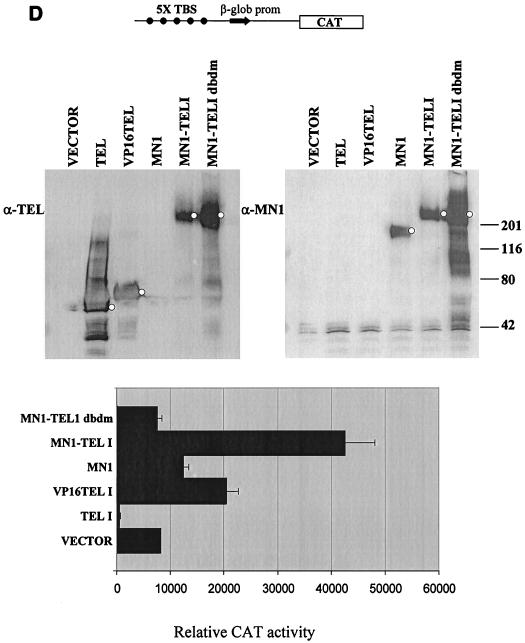
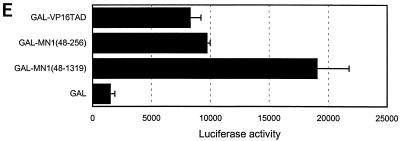
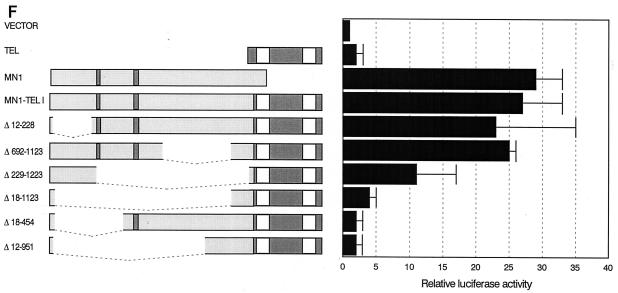
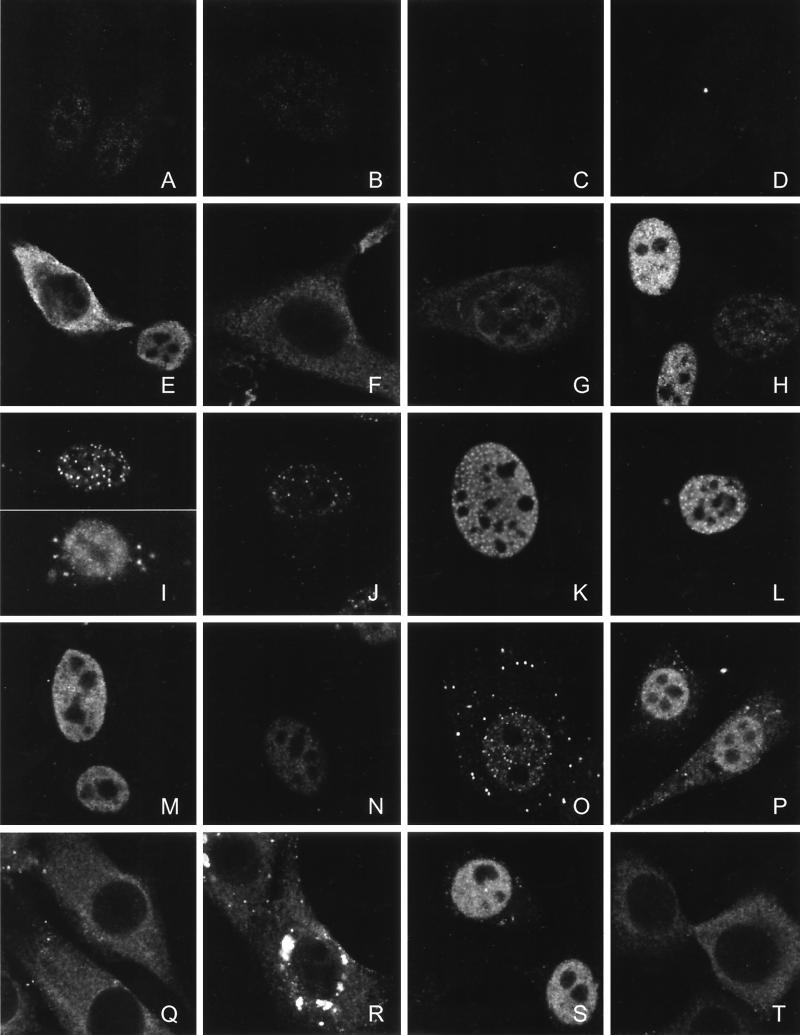
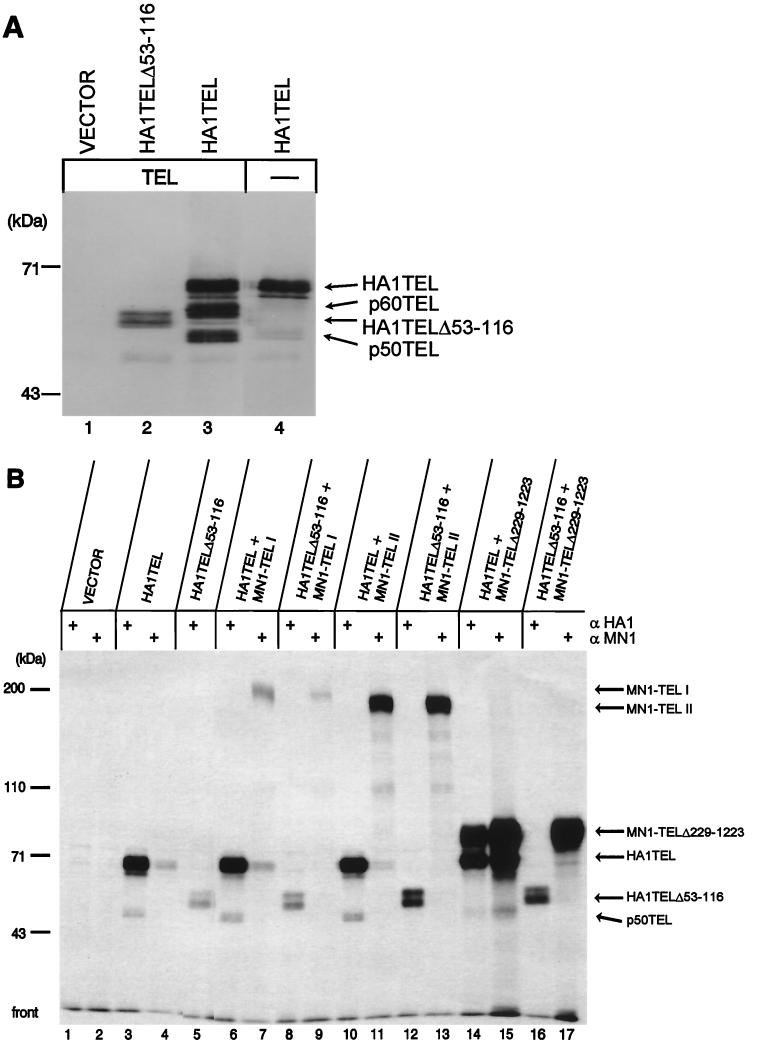
The Tel gene (or ETV6) is the target of the translocation (12;22)(p13;q11) in myeloid leukemia. TEL is a member of the ETS family of transcription factors and contains the pointed protein interaction (PNT) domain and an ETS DNA binding domain (DBD). By contrast to other chimeric proteins that contain TEL's PNT domain, such as TEL-platelet-derived growth factor beta receptor in t(5;12)(q33;p13), MN1-TEL contains the DBD of TEL. The N-terminal MN1 moiety is rich in proline residues and contains two polyglutamine stretches, suggesting that MN1-TEL may act as a deregulated transcription factor. We now show that MN1-TEL type I, unlike TEL and MN1, transforms NIH 3T3 cells. The transforming potential depends on both N-terminal MN1 sequences and a functional TEL DBD. Furthermore, we demonstrate that MN1 has transcription activity and that MN1-TEL acts as a chimeric transcription factor on the Moloney sarcoma virus long terminal repeat and a synthetic promoter containing TEL binding sites. The transactivating capacity of MN1-TEL depended on both the DBD of TEL and sequences in MN1. MN1-TEL contributes to leukemogenesis by a mechanism distinct from that of other chimeric proteins containing TEL.
Figures
References
-
- Buijs A, Sherr S, van Baal S, van Bezouw S, van der Plas D, Geurts van Kessel A, Riegman P, Lekanne Deprez R, Zwarthoff E, Hagemeijer A, Grosveld G. Translocation (12;22)(p13;q11) in myeloproliferative disorders results in fusion of the ETS-like TEL gene on 12p13 to the MN1 gene on 22q11. Oncogene. 1995;10:1511–1519. - PubMed
-
- Carroll M, Tomasson M H, Barker G F, Golub T R, Gilliland D G. The TEL/platelet-derived growth factor β receptor (PDGFβR) fusion in chronic myelomonocytic leukemia is a transforming protein that self-associates and activates PDGFβR kinase-dependent signaling pathways. Proc Natl Acad Sci USA. 1996;93:14845–14850. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources