

Components and dynamics of fiber formation define a ubiquitous biogenesis pathway for bacterial pili
- PMID: 11101514
- PMCID: PMC305860
- DOI: 10.1093/emboj/19.23.6408
Components and dynamics of fiber formation define a ubiquitous biogenesis pathway for bacterial pili
Abstract
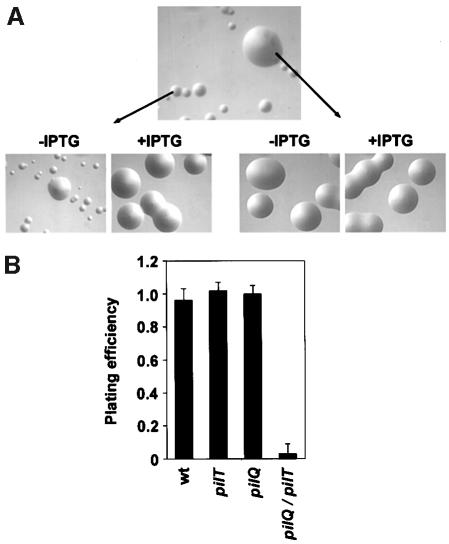
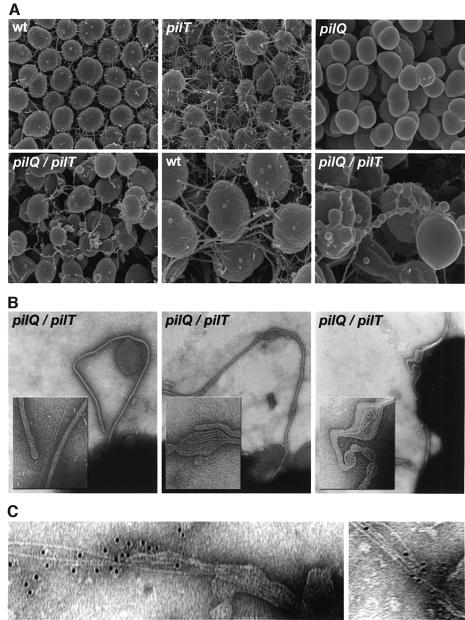
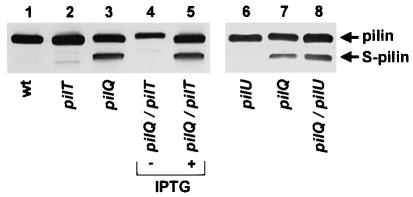
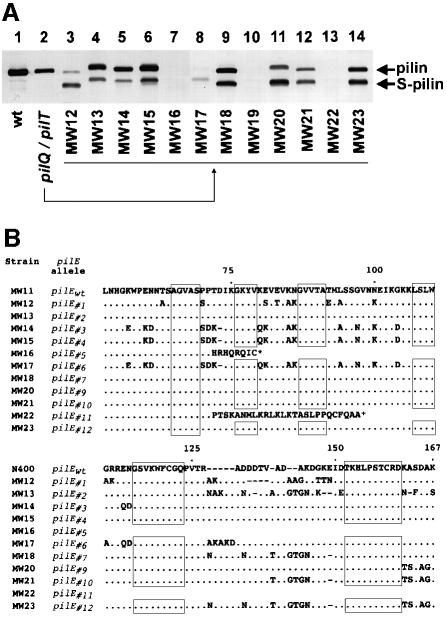
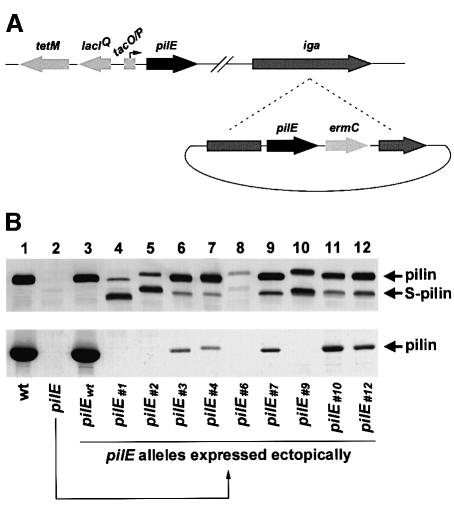
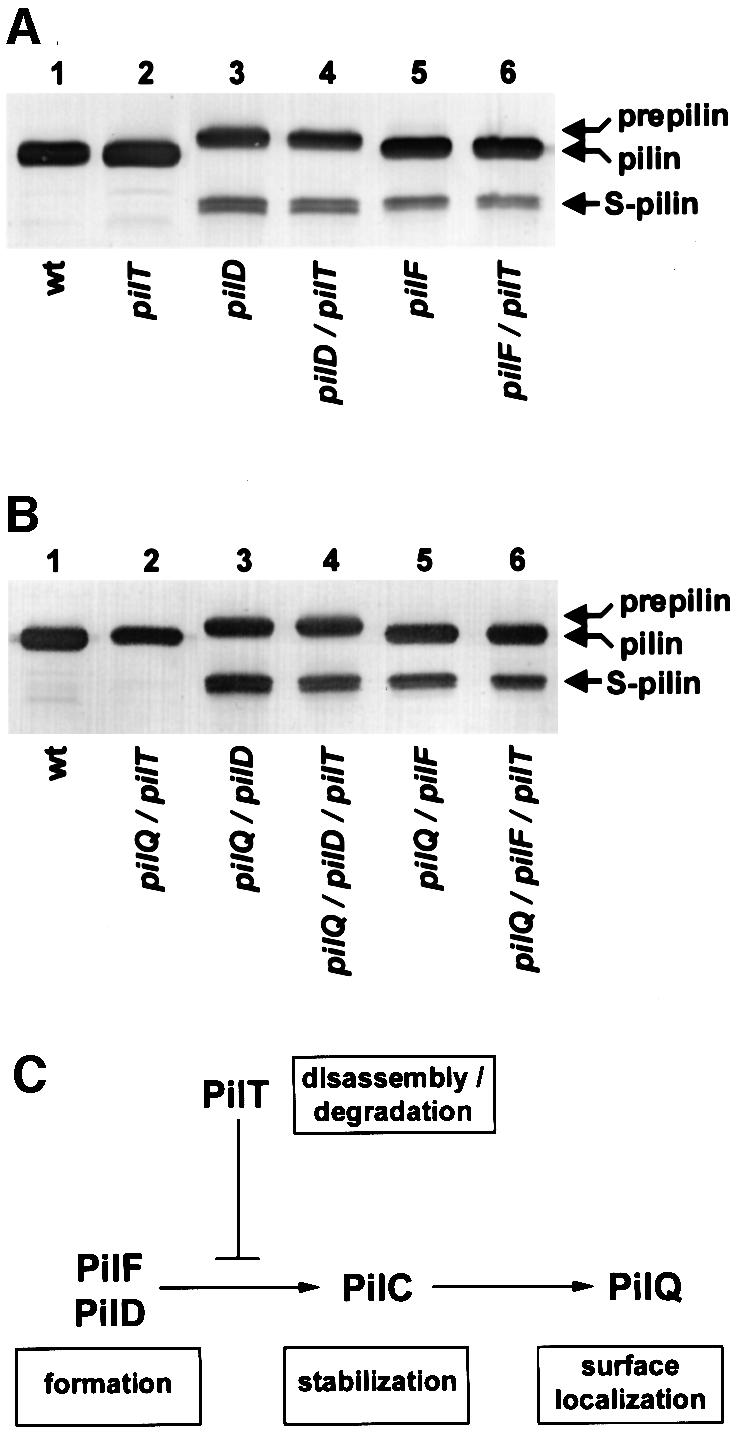
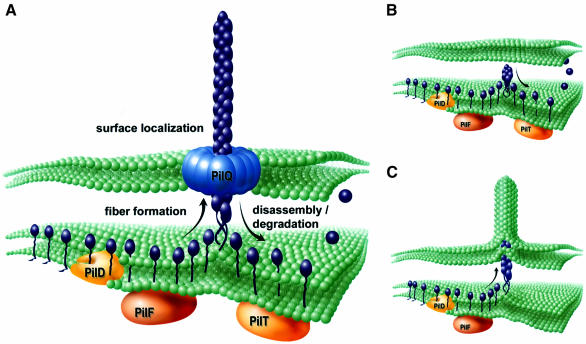
Type IV pili (Tfp) are a unique class of multifunctional surface organelles in Gram-negative bacteria, which play important roles in prokaryotic cell biology. Although components of the Tfp biogenesis machinery have been characterized, it is not clear how they function or interact. Using Neisseria gonorrhoeae as a model system, we report here that organelle biogenesis can be resolved into two discrete steps: fiber formation and translocation of the fiber to the cell surface. This conclusion is based on the capturing of an intermediate state in which the organelle is retained within the cell owing to the simultaneous absence of the secretin family member and biogenesis component PilQ and the twitching motility/pilus retraction protein PilT. This finding is the first demonstration of a specific translocation defect associated with loss of secretin function, and additionally confirms the role of PilT as a conditional antagonist of stable pilus fiber formation. These findings have important implications for Tfp structure and function and are pertinent to other membrane translocation systems that utilize a highly related set of components.
Figures
References
-
- Bitter W., Koster,M., Latijnhouwers,M., de Cock,H. and Tommassen,J. (1998) Formation of oligomeric rings by XcpQ and PilQ, which are involved in protein transport across the outer membrane of Pseudomonas aeruginosa. Mol. Microbiol., 27, 209–219. - PubMed
-
- Bradley D.E. (1980) A function of Pseudomonas aeruginosa PAO polar pili: twitching motility. Can. J. Microbiol., 26, 146–154. - PubMed
-
- Crago A.M. and Koronakis,V. (1998) Salmonella InvG forms a ring-like multimer that requires the InvH lipoprotein for outer membrane localization. Mol. Microbiol., 30, 47–56. - PubMed
-
- Dougherty B.A. and Smith,H.O. (1999) Identification of Haemophilus influenzae Rd transformation genes using cassette mutagenesis. Microbiology, 145, 401–409. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
