

Enhanced vulnerability to cocaine self-administration is associated with elevated impulse activity of midbrain dopamine neurons
- PMID: 11102497
- PMCID: PMC6773051
- DOI: 10.1523/JNEUROSCI.20-23-08876.2000
Enhanced vulnerability to cocaine self-administration is associated with elevated impulse activity of midbrain dopamine neurons
Abstract
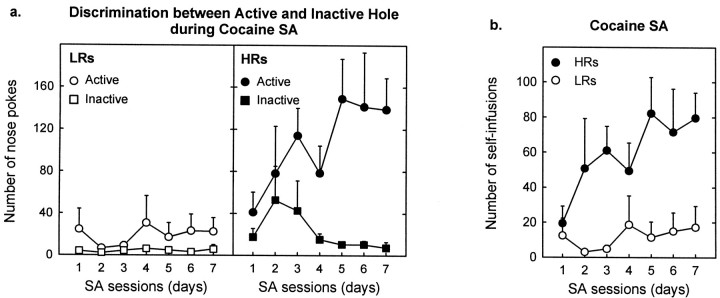
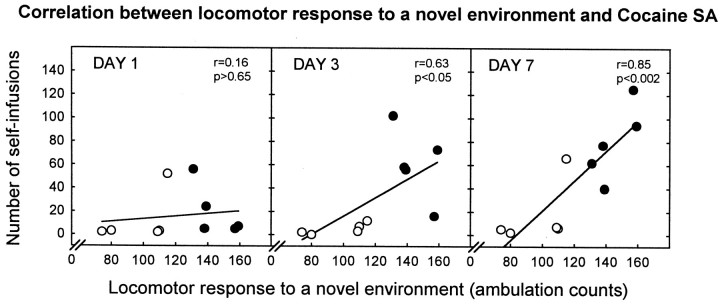
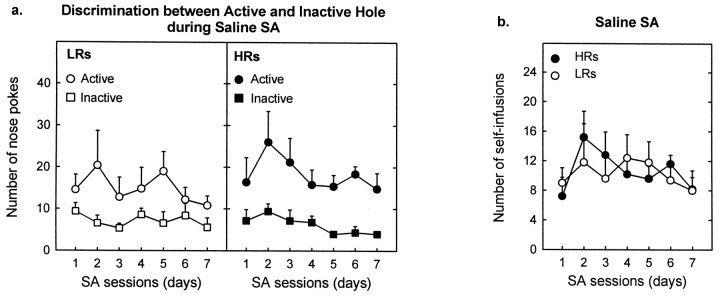
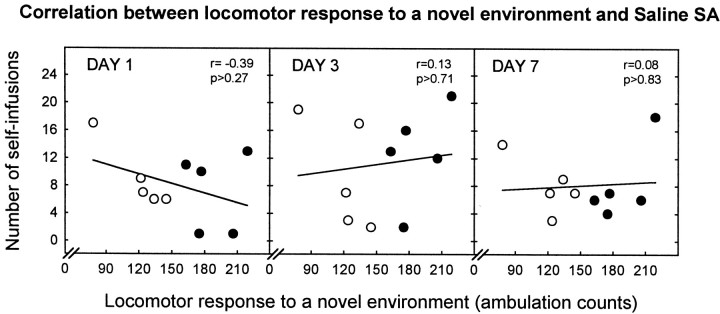
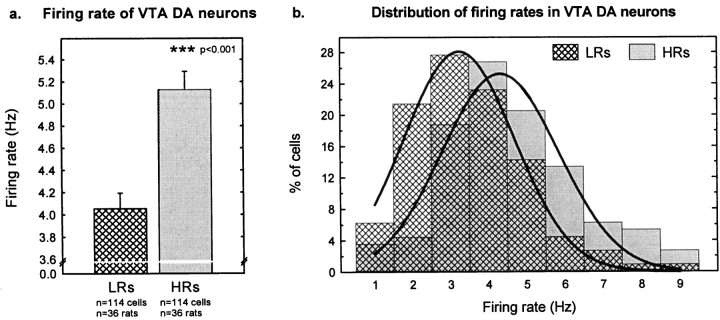
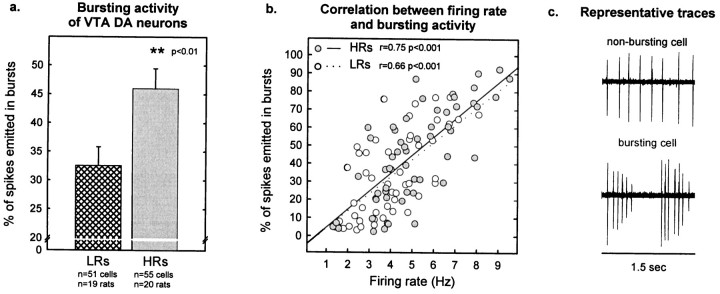
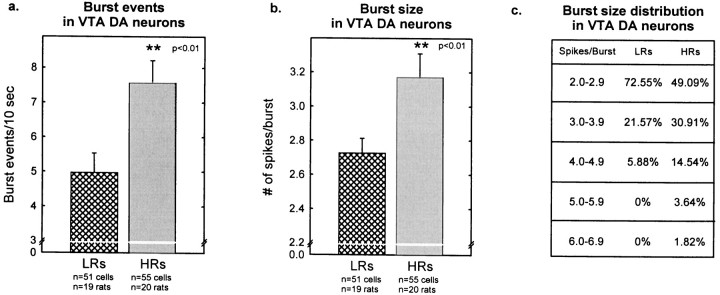
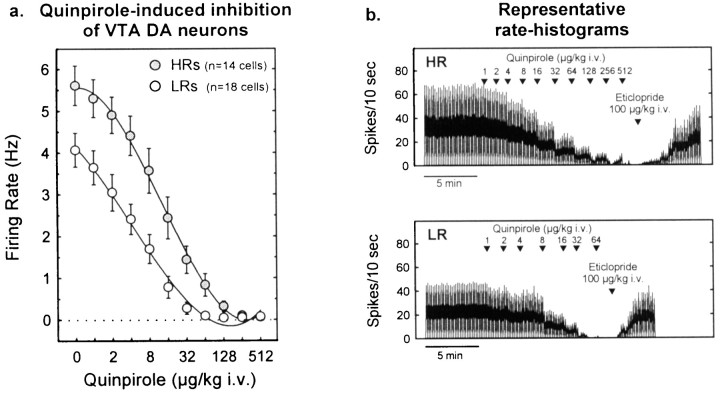
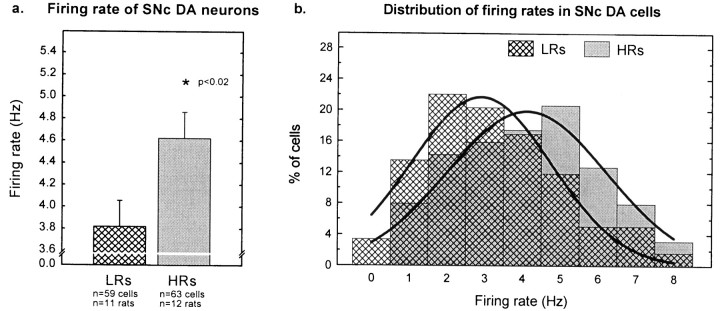
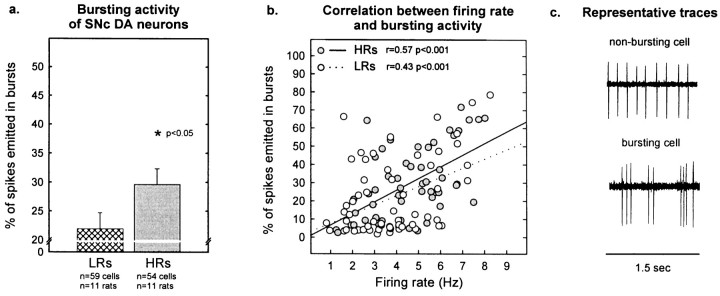
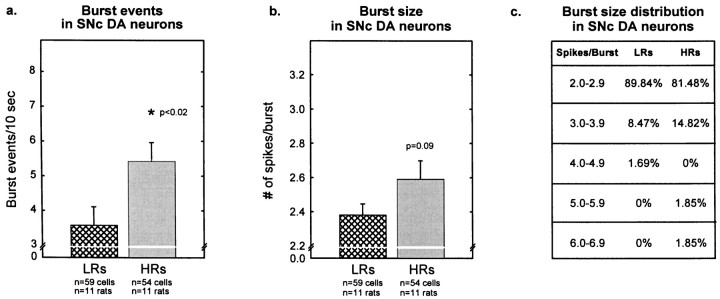
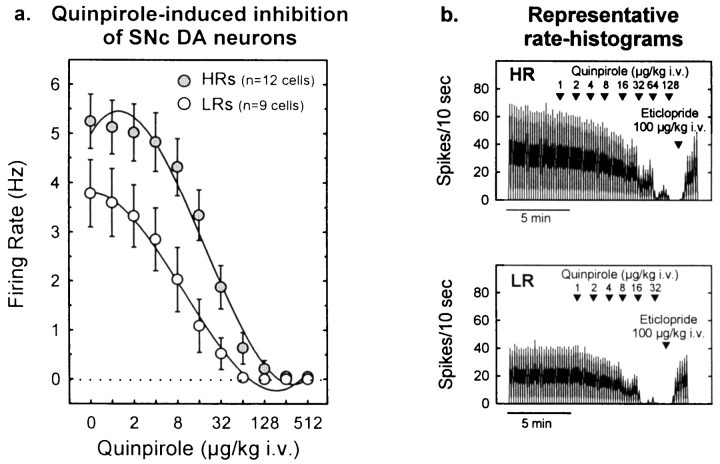
Individual differences in responding to a novel environment predict behavioral and neurochemical responses to psychostimulant drugs. Rats with a high locomotor response to a novel environment (HRs) exhibit enhanced self-administration (SA) behavior, sensitization, and basal or drug-induced dopamine release in the nucleus accumbens compared with rats with a low response to the novel context (LRs). In this study, we determined whether such differences in vulnerability to drug addiction might be related to differences in dopamine (DA) neuron activity. Rats were divided into HRs and LRs according to their response to a novel environment and then tested for acquisition of cocaine SA. HRs rapidly acquired cocaine SA (175 microg/kg per infusion), whereas LRs did not. Differences in cocaine SA were not caused by differences in exploratory behavior or sampling because these behaviors did not differ in HRs and LRs self-administering a saline solution. In a separate experiment, we used extracellular single-unit recordings and found that HRs exhibit higher basal firing rates and bursting activity of DA neurons in the ventral tegmental area and, to a lesser extent, in the substantia nigra pars compacta. The greater activity of midbrain DA cells in HRs was accompanied by reduced sensitivity to the inhibitory effects of a DA D2-class receptor agonist, indicating possible subsensitivity of impulse-regulating DA autoreceptors. These results demonstrate that differences in the basal activity of DA neurons may be critically involved in determining individual vulnerability to drugs of abuse.
Figures
References
-
- Ackerman JM, White FJ. A10 somatodendritic dopamine autoreceptor sensitivity following withdrawal from repeated cocaine treatment. Neurosci Lett. 1990;117:181–187. - PubMed
-
- Ackerman JM, White FJ. Decreased activity of rat A10 dopamine neurons following withdrawal from repeated cocaine. Eur J Pharmacol. 1992;218:171–173. - PubMed
-
- Aghajanian GK, Bunney BS. Pharmacological characterization of dopamine “autoreceptors” by microiontophoretic single-cell recording studies. Adv Biochem Psychopharmacol. 1977a;16:433–438. - PubMed
-
- Aghajanian GK, Bunney BS. Dopamine “autoreceptors”: pharmacological characterization by microiontophoretic single cell recording studies. Naunyn Schmiedebergs Arch Pharmacol. 1977b;297:1–7. - PubMed
-
- Amini B, Clark JW, Jr, Canavier CC. Calcium dynamics underlying pacemaker-like and burst firing oscillations in midbrain dopaminergic neurons: a computational study. J Neurophysiol. 1999;82:2249–2261. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous