

Astroglial expression of human alpha(1)-antichymotrypsin enhances alzheimer-like pathology in amyloid protein precursor transgenic mice
- PMID: 11106573
- PMCID: PMC1885780
- DOI: 10.1016/s0002-9440(10)64839-0
Astroglial expression of human alpha(1)-antichymotrypsin enhances alzheimer-like pathology in amyloid protein precursor transgenic mice
Abstract
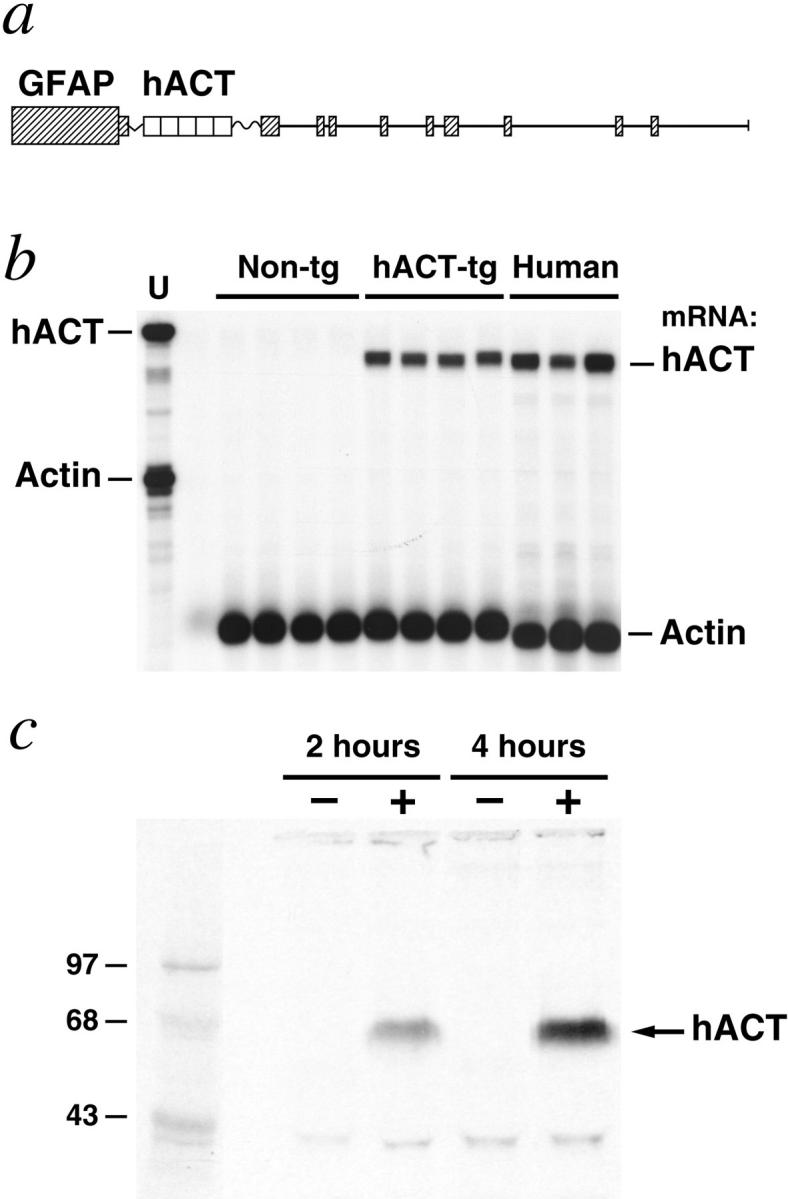
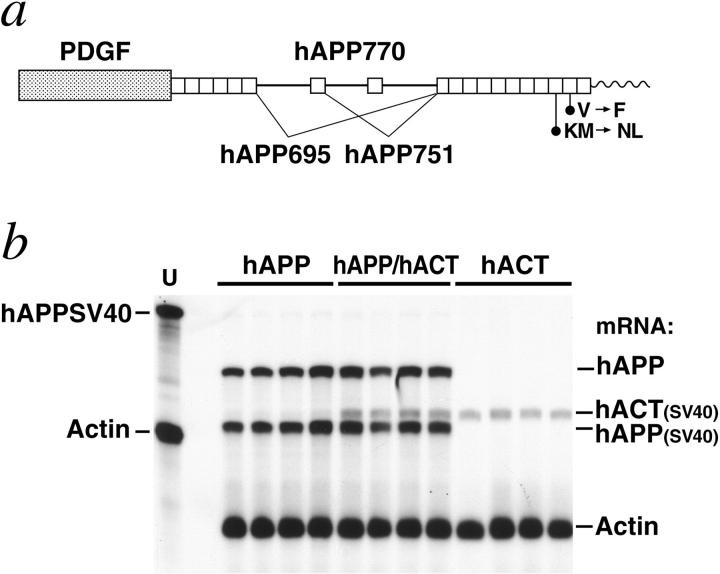
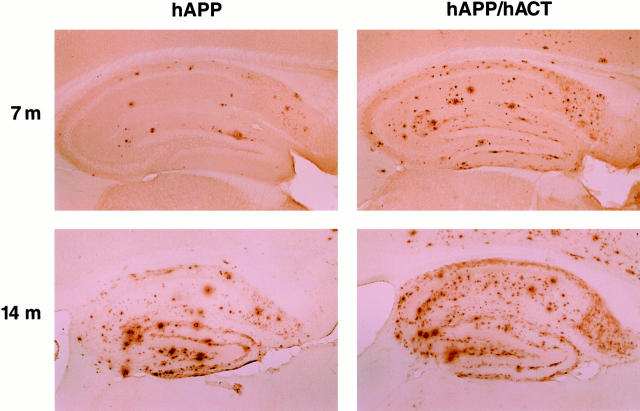
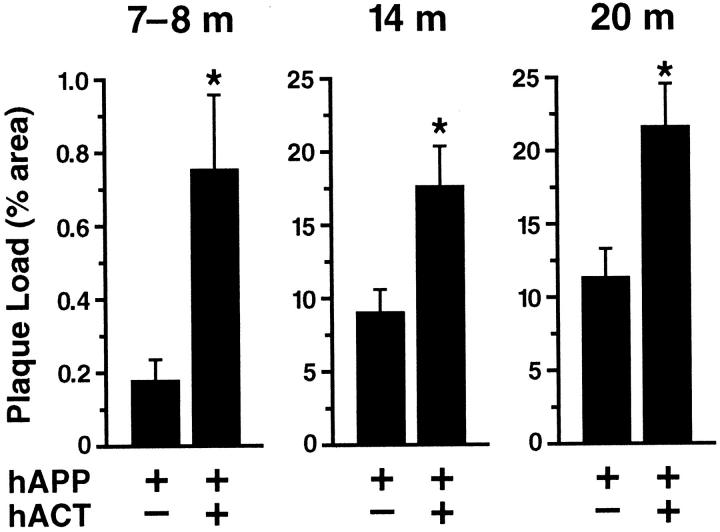
Proteases and their inhibitors play key roles in physiological and pathological processes. Cerebral amyloid plaques are a pathological hallmark of Alzheimer's disease (AD). They contain amyloid-ss (Ass) peptides in tight association with the serine protease inhibitor alpha(1)-antichymotrypsin.(1,2) However, it is unknown whether the increased expression of alpha(1)-antichymotrypsin found in AD brains counteracts or contributes to the disease. We used regulatory sequences of the glial fibrillary acidic protein gene(3) to express human alpha(1)-antichymotrypsin (hACT) in astrocytes of transgenic mice. These mice were crossed with transgenic mice that produce human amyloid protein precursors (hAPP) and Ass in neurons.(4,5) No amyloid plaques were found in transgenic mice expressing hACT alone, whereas hAPP transgenic mice and hAPP/hACT doubly transgenic mice developed typical AD-like amyloid plaques in the hippocampus and neocortex around 6 to 8 months of age. Co-expression of hAPP and hACT significantly increased the plaque burden at 7 to 8, 14, and 20 months. Both hAPP and hAPP/hACT mice showed significant decreases in synaptophysin-immunoreactive presynaptic terminals in the dentate gyrus, compared with nontransgenic littermates. Our results demonstrate that hACT acts as an amyloidogenic co-factor in vivo and suggest that the role of hACT in AD is pathogenic.
Figures

References
-
- Abraham CR, Selkoe DJ, Potter H: Immunochemical identification of the serine protease inhibitor α1-antichymotrypsin in the brain amyloid deposits of Alzheimer’s disease. Cell 1988, 52:487-501 - PubMed
-
- Johnson WB, Ruppe MD, Rockenstein EM, Price J, Sarthy VP, Verderber LC, Mucke L: Indicator expression directed by regulatory sequences of the glial fibrillary acidic protein (GFAP) gene: in vivo comparison of distinct GFAP-lacZ transgenes. Glia 1995, 13:174-184 - PubMed
-
- Mucke L, Masliah E, Yu G-Q, Mallory M, Rockenstein EM, Tatsuno G, Hu K, Kholodenko D, Johnson-Wood K, McConlogue L: High-level neuronal expression of Aβ1-42 in wild-type human amyloid protein precursor transgenic mice: synaptotoxicity without plaque formation. J Neurosci 2000, 20:4050-4058 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
