

Defining roles for HOX and MEIS1 genes in induction of acute myeloid leukemia
- PMID: 11113197
- PMCID: PMC88796
- DOI: 10.1128/MCB.21.1.224-234.2001
Defining roles for HOX and MEIS1 genes in induction of acute myeloid leukemia
Abstract
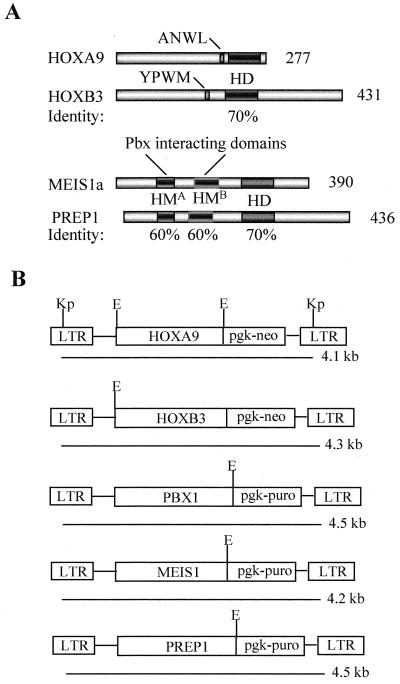
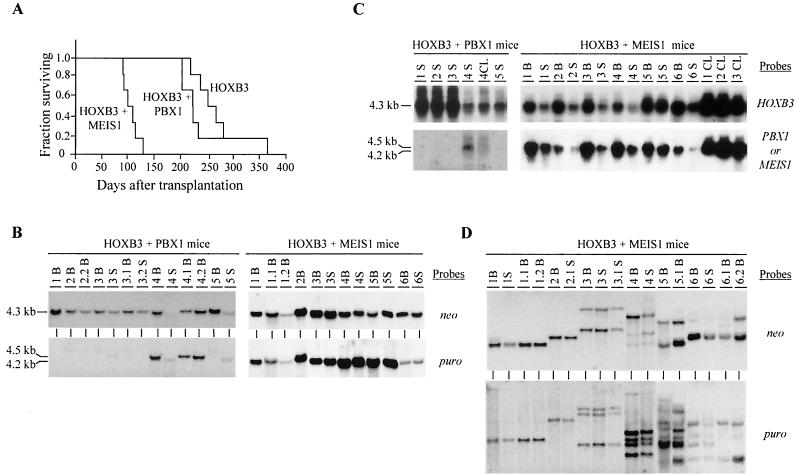
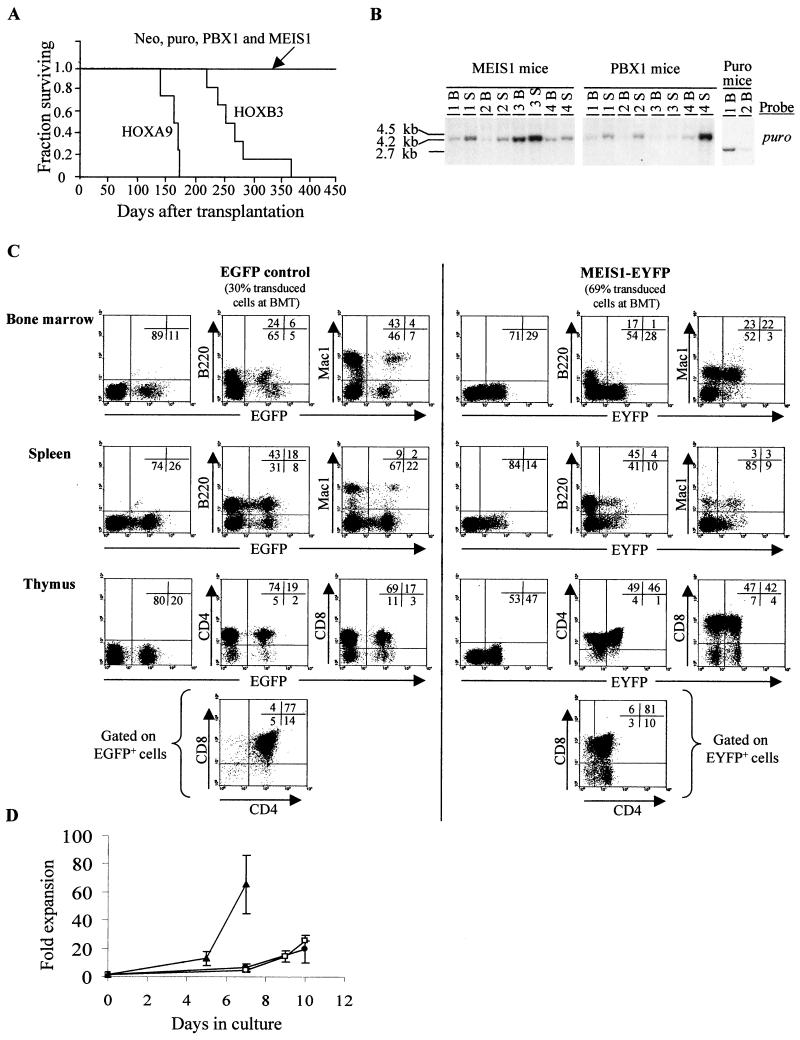
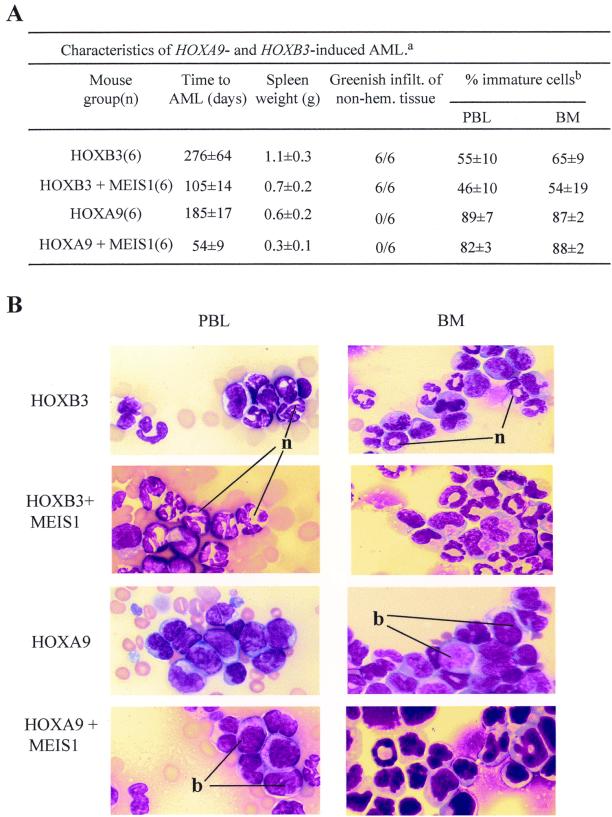
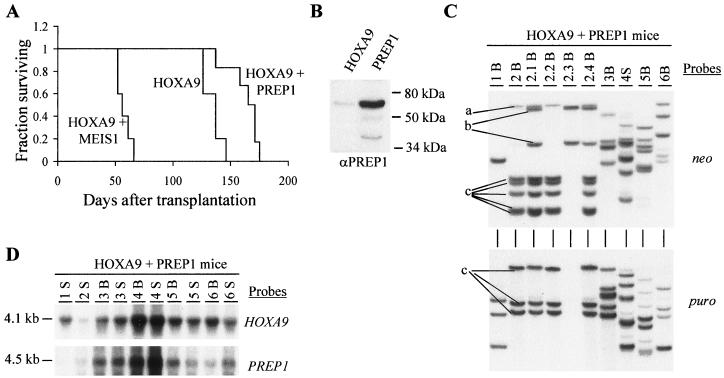
Complex genetic and biochemical interactions between HOX proteins and members of the TALE (i.e., PBX and MEIS) family have been identified in embryonic development, and some of these interactions also appear to be important for leukemic transformation. We have previously shown that HOXA9 collaborates with MEIS1 in the induction of acute myeloid leukemia (AML). In this report, we demonstrate that HOXB3, which is highly divergent from HOXA9, also genetically interacts with MEIS1, but not with PBX1, in generating AML. In addition, we show that the HOXA9 and HOXB3 genes play key roles in establishing all the main characteristics of the leukemias, while MEIS1 functions only to accelerate the onset of the leukemic transformation. Contrasting the reported functional similarities between PREP1 and MEIS1, such as PBX nuclear retention, we also show that PREP1 overexpression is incapable of accelerating the HOXA9-induced AML, suggesting that MEIS1 function in transformation must entail more than PBX nuclear localization. Collectively, these data demonstrate that MEIS1 is a common leukemic collaborator with two structurally and functionally divergent HOX genes and that, in this collaboration, the HOX gene defines the identity of the leukemia.
Figures
Similar articles
-
Hoxa9 transforms primary bone marrow cells through specific collaboration with Meis1a but not Pbx1b.EMBO J. 1998 Jul 1;17(13):3714-25. doi: 10.1093/emboj/17.13.3714. EMBO J. 1998. PMID: 9649441 Free PMC article.
-
An inhibitory switch derepressed by pbx, hox, and Meis/Prep1 partners regulates DNA-binding by pbx1 and E2a-pbx1 and is dispensable for myeloid immortalization by E2a-pbx1.Oncogene. 1999 Dec 23;18(56):8033-43. doi: 10.1038/sj.onc.1203377. Oncogene. 1999. PMID: 10637514
-
Persistent transactivation by meis1 replaces hox function in myeloid leukemogenesis models: evidence for co-occupancy of meis1-pbx and hox-pbx complexes on promoters of leukemia-associated genes.Mol Cell Biol. 2006 May;26(10):3902-16. doi: 10.1128/MCB.26.10.3902-3916.2006. Mol Cell Biol. 2006. PMID: 16648484 Free PMC article.
-
A tale of TALE, PREP1, PBX1, and MEIS1: Interconnections and competition in cancer.Bioessays. 2017 May;39(5). doi: 10.1002/bies.201600245. Epub 2017 Mar 21. Bioessays. 2017. PMID: 28322463 Review.
-
The Historical Relationship Between Meis1 and Leukemia.Adv Exp Med Biol. 2022;1387:127-144. doi: 10.1007/5584_2021_705. Adv Exp Med Biol. 2022. PMID: 35304708 Review.
Cited by
-
Differential transcriptional regulation of meis1 by Gfi1b and its co-factors LSD1 and CoREST.PLoS One. 2013;8(1):e53666. doi: 10.1371/journal.pone.0053666. Epub 2013 Jan 7. PLoS One. 2013. PMID: 23308270 Free PMC article.
-
Up-regulation of a HOXA-PBX3 homeobox-gene signature following down-regulation of miR-181 is associated with adverse prognosis in patients with cytogenetically abnormal AML.Blood. 2012 Mar 8;119(10):2314-24. doi: 10.1182/blood-2011-10-386235. Epub 2012 Jan 17. Blood. 2012. PMID: 22251480 Free PMC article.
-
Multiple machine-learning tools identifying prognostic biomarkers for acute Myeloid Leukemia.BMC Med Inform Decis Mak. 2024 Jan 2;24(1):2. doi: 10.1186/s12911-023-02408-9. BMC Med Inform Decis Mak. 2024. PMID: 38167056 Free PMC article.
-
The Critical Role of TRIB2 in Cancer and Therapy Resistance.Cancers (Basel). 2021 May 30;13(11):2701. doi: 10.3390/cancers13112701. Cancers (Basel). 2021. PMID: 34070799 Free PMC article. Review.
-
NUP98-HOXA9 expression in hemopoietic stem cells induces chronic and acute myeloid leukemias in mice.EMBO J. 2001 Feb 1;20(3):350-61. doi: 10.1093/emboj/20.3.350. EMBO J. 2001. PMID: 11157742 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources