

Negative regulation of defense responses in plants by a conserved MAPKK kinase
- PMID: 11114160
- PMCID: PMC14597
- DOI: 10.1073/pnas.98.1.373
Negative regulation of defense responses in plants by a conserved MAPKK kinase
Abstract
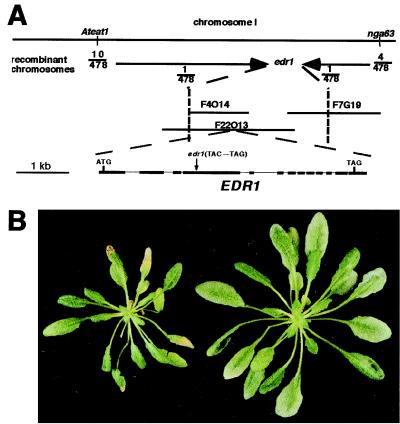
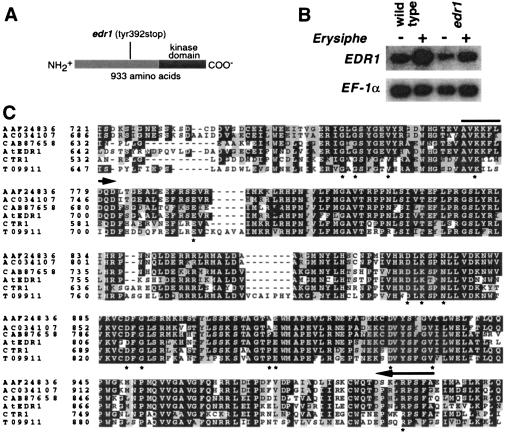
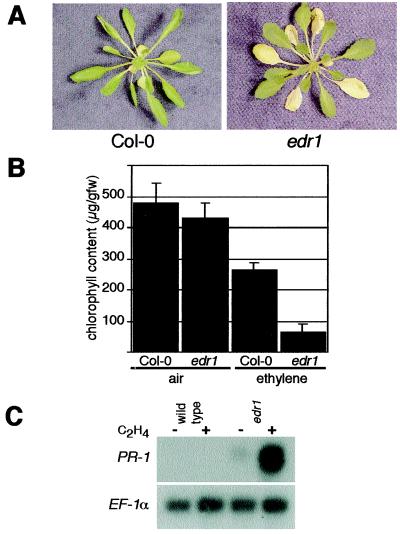
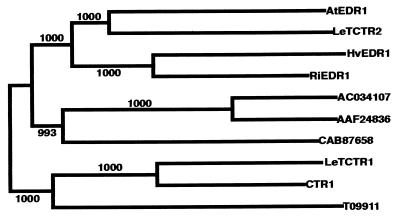
The enhanced disease resistance 1 (edr1) mutation of Arabidopsis confers resistance to powdery mildew disease caused by the fungus Erysiphe cichoracearum. Resistance mediated by the edr1 mutation is correlated with induction of several defense responses, including host cell death. Double mutant analysis revealed that all edr1-associated phenotypes are suppressed by mutations that block salicylic acid (SA) perception (nim1) or reduce SA production (pad4 and eds1). The NahG transgene, which lowers endogenous SA levels, also suppressed edr1. In contrast, the ein2 mutation did not suppress edr1-mediated resistance and associated phenotypes, indicating that ethylene and jasmonic acid-induced responses are not required for edr1 resistance. The EDR1 gene was isolated by positional cloning and was found to encode a putative MAP kinase kinase kinase similar to CTR1, a negative regulator of ethylene responses in Arabidopsis. Taken together, these data suggest that EDR1 functions at the top of a MAP kinase cascade that negatively regulates SA-inducible defense responses. Putative orthologs of EDR1 are present in monocots such as rice and barley, indicating that EDR1 may regulate defense responses in a wide range of crop species.
Figures

Comment in
-
Plant mitogen-activated protein kinase cascades: Negative regulatory roles turn out positive.Proc Natl Acad Sci U S A. 2001 Jan 30;98(3):784-6. doi: 10.1073/pnas.98.3.784. Proc Natl Acad Sci U S A. 2001. PMID: 11158543 Free PMC article. Review. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
