

Review
doi: 10.1016/s0166-3542(00)00126-1.
Innate antiviral defenses in body fluids and tissues
Affiliations
- PMID: 11114410
- PMCID: PMC7125796
- DOI: 10.1016/s0166-3542(00)00126-1
Item in Clipboard
Review
Innate antiviral defenses in body fluids and tissues
Antiviral Res.
2000 Nov.
Abstract
Innate, non-specific, resistance mechanisms are important barriers to pathogens, particularly delaying virus multiplication at the onset of infections. These innate defense mechanisms include a series of mechanical barriers, pre-existing inhibitory molecules, and cellular responses with antimicrobial activity. The antiviral activities of these innate inhibitors reside in a variety of partly characterized substances. This review presents the innate antiviral inhibitors in cell cultures, urine, serum, the gastrointestinal tract, the nervous system, tissues of crustaceans, and saliva. Medical adaptation of the innate antiviral defense mechanisms may be useful for prevention and treatment of viral infections.
Figures
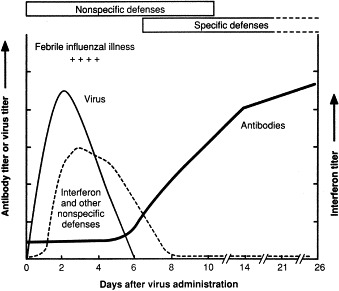
Production of virus, interferon, and antibody during experimental infection of humans with influenza wild-type virus. Non-specific defenses include anatomic barriers, inhibitors, phagocytosis, fever, inflammation and IFN. Specific defenses include antibody and cell-mediated immunity. Data from a study by B. Murphy et al., National Institutes of Health (pers. commun.).
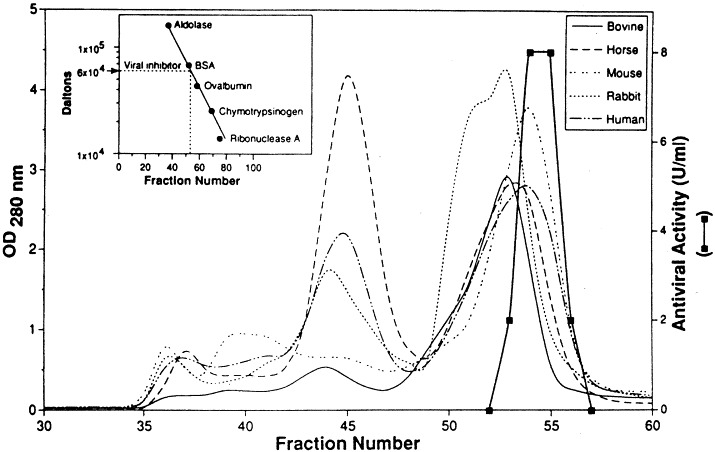
Estimation of the molecular size of the UTIβ by size exclusion chromatography. Sera from different mammalian species were individually loaded onto a column (2.5×90 cm) of Sephacryl S-200. The column was equilibrated with phosphate buffered saline. Four-milliliter fractions were collected: the antiviral activity (depicted by the heavy line) in individual fractions was examined by the standard plaque reduction assay. The inset shows the calibration of the column. Reprinted with permission from the J. Biol. Regul. Homeost. Agents, 1993, 7: 9.
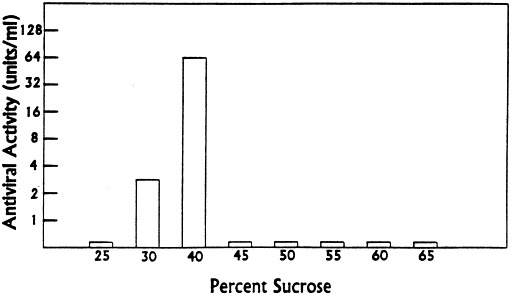
Sedimentation rate of virus inhibitor from lamb brain tissue in a discontinuous sucrose gradient. Sucrose solutions in PBS, 5 mM MgC12, were used to form step gradients. Equivalent volumes of brain inhibitor preparation (0.5 ml) and 50% sucrose were mixed, layered on the discontinuous gradient, and centrifuged at 31 000 rev./min in a SW-55 rotor for 90 min. Collected fractions were examined for antiviral activity against Sindbis virus after extensive dialysis. Reprinted with permission from Antiviral Res., 1995, 27:380.
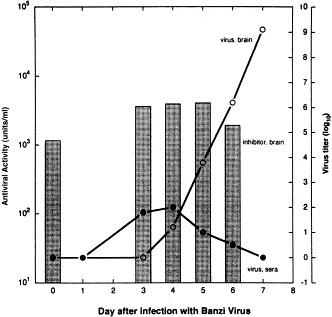
Level of virus inhibitor in mouse brain extracts is unchanged during encephalitis. Three-week-old female outbred (ICR) mice were infected intraperitoneally with 3 LD75 of BZV. Brains were harvested at days 3–6 postinfection, and homogenized. Virus inhibitory levels in the homogenates were titered against Sindbis virus, and are depicted in the bar graph (left Y axis). The rise in the virus titers in the brains and sera of infected mice (taken from Singh et al., 1989) are superimposed to illustrate the course of the BZV infections (○, virus load in brain; •, virus load per ml serum). Reprinted with permission from Antiviral Res., 1995, 27:384.
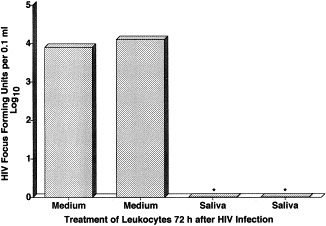
Saliva interrupts the multiplication of human immunodeficiency virus (HIV) in infected human mononuclear leukocytes. *P<0.05 by Student's t-test. Reprinted with permission from Arch. Intern. Med., 1999, 159:306.
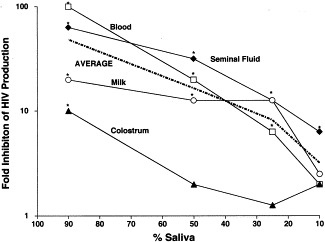
Dilution in blood, milk, colostrums, and seminal fluid prevents saliva's inhibition of HIV multiplication in human PBL. *P<0.05 Student's t-test. Reprinted with permission from J. Infect. Dis., 2000, 181:500.
Similar articles
-
Broad antiviral activity in tissues of crustaceans.Antiviral Res. 2000 Oct;48(1):39-47. doi: 10.1016/s0166-3542(00)00117-0. Antiviral Res. 2000. PMID: 11080539
-
HIV Exploits Antiviral Host Innate GCN2-ATF4 Signaling for Establishing Viral Replication Early in Infection.mBio. 2017 May 2;8(3):e01518-16. doi: 10.1128/mBio.01518-16. mBio. 2017. PMID: 28465428 Free PMC article.
-
Antiviral immunity in crustaceans.Fish Shellfish Immunol. 2009 Aug;27(2):79-88. doi: 10.1016/j.fsi.2009.02.009. Epub 2009 Feb 15. Fish Shellfish Immunol. 2009. PMID: 19223016 Free PMC article. Review.
-
[Advances in Avian Antiviral Innate Immune Effectors].Bing Du Xue Bao. 2016 Sep;32(5):627-33. Bing Du Xue Bao. 2016. PMID: 30003769 Review. Chinese.
-
Mitophagy in viral infections.DNA Cell Biol. 2014 Nov;33(11):739-42. doi: 10.1089/dna.2014.2567. Epub 2014 Jul 22. DNA Cell Biol. 2014. PMID: 25050805
Cited by
-
Activity and regulation of alpha interferon in respiratory syncytial virus and human metapneumovirus experimental infections.J Virol. 2005 Aug;79(16):10190-9. doi: 10.1128/JVI.79.16.10190-10199.2005. J Virol. 2005. PMID: 16051812 Free PMC article.
-
Development and validation of an assay to detect porcine reproductive and respiratory syndrome virus-specific neutralizing antibody titers in pig oral fluid samples.Clin Vaccine Immunol. 2013 Aug;20(8):1305-13. doi: 10.1128/CVI.00276-13. Epub 2013 Jun 19. Clin Vaccine Immunol. 2013. PMID: 23784856 Free PMC article.
-
Respiratory syncytial virus infection induces higher Toll-like receptor-3 expression and TNF-α production than human metapneumovirus infection.PLoS One. 2013 Sep 9;8(9):e73488. doi: 10.1371/journal.pone.0073488. eCollection 2013. PLoS One. 2013. PMID: 24039959 Free PMC article.
-
Fellatio by fruit bats prolongs copulation time.PLoS One. 2009 Oct 28;4(10):e7595. doi: 10.1371/journal.pone.0007595. PLoS One. 2009. PMID: 19862320 Free PMC article.
References
-
- Alpers D.H. Uptake and fate of absorbed amino acids and peptides in the mammalian intestine. Fed. Proc. 1986;45:2261–2267. - PubMed
-
- Archibald D.W., Cole G.A. In vitro inhibition of HIV-1 infectivity by human salivas. AIDS Res. Hum. Retroviruses. 1990;6:1425–1432. - PubMed
-
- Banfield W.J., Kisch A.L. The effect of chloroquine on herpesvirus infection in vitro and in vivo. Proc. Soc. Exp. Biol. Med. 1973;142:1018–1022. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
