

FtsK functions in the processing of a Holliday junction intermediate during bacterial chromosome segregation
- PMID: 11114887
- PMCID: PMC317095
- DOI: 10.1101/gad.188700
FtsK functions in the processing of a Holliday junction intermediate during bacterial chromosome segregation
Abstract
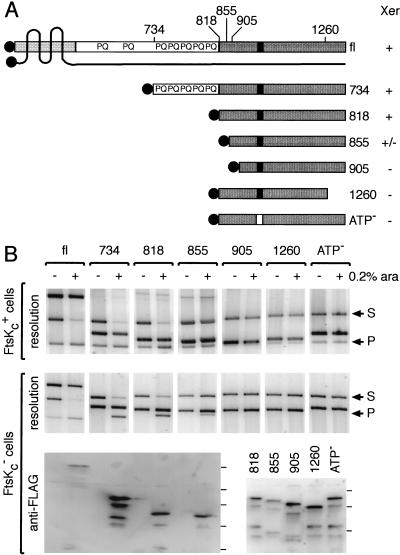
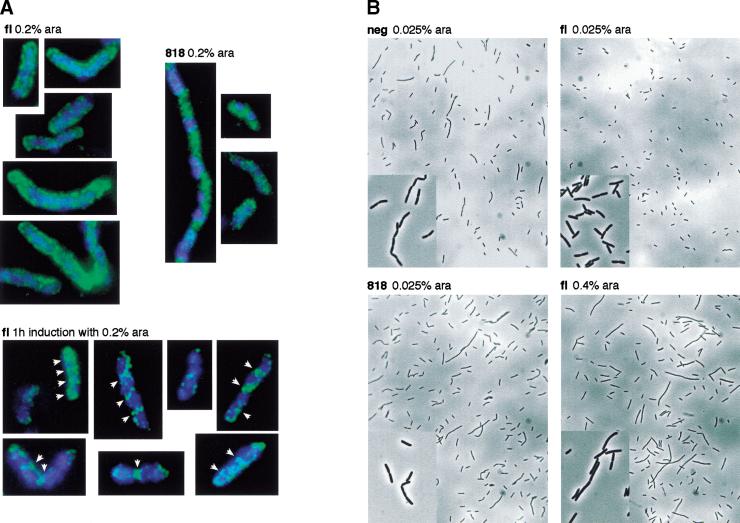
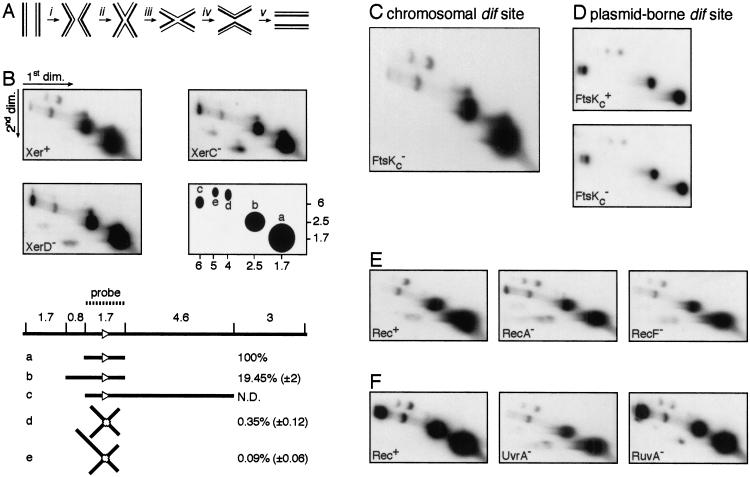
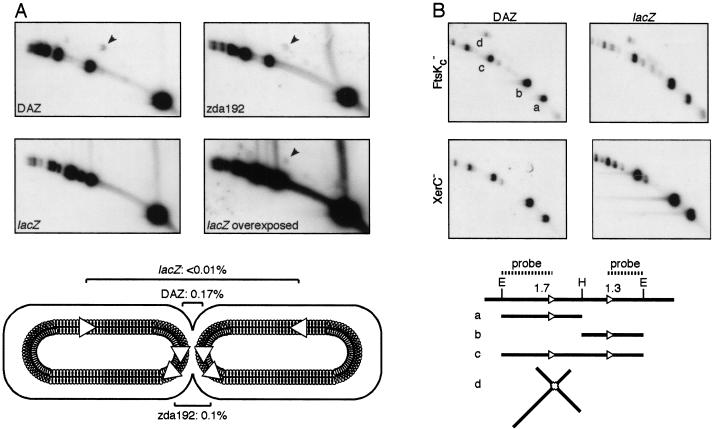
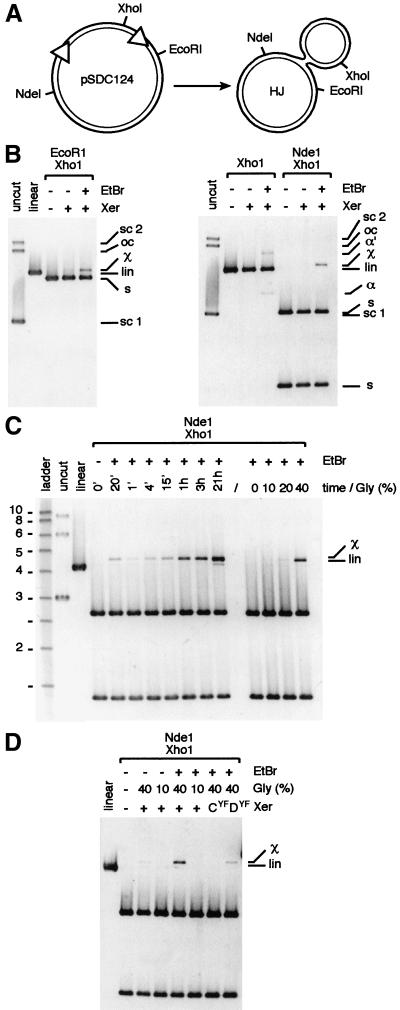
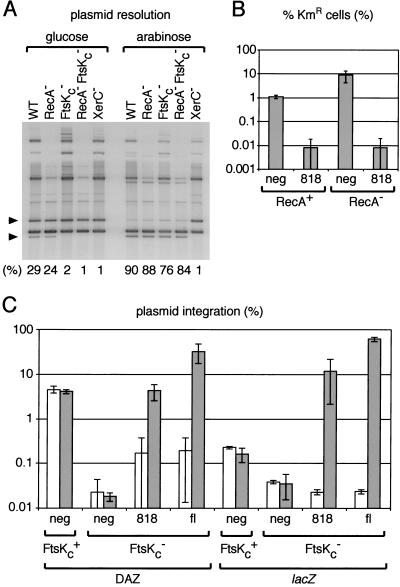
In bacteria with circular chromosomes, homologous recombination can generate chromosome dimers that cannot be segregated to daughter cells at cell division. Xer site-specific recombination at dif, a 28-bp site located in the replication terminus region of the chromosome, converts dimers to monomers through the sequential action of the XerC and XerD recombinases. Chromosome dimer resolution requires that dif is positioned correctly in the chromosome, and the activity of FtsK, a septum-located protein that coordinates cell division with chromosome segregation. Here, we show that cycles of XerC-mediated strand exchanges form and resolve Holliday junction intermediates back to substrate irrespective of whether conditions support a complete recombination reaction. The C-terminal domain of FtsK is sufficient to activate the exchange of the second pair of strands by XerD, allowing both intra- and intermolecular recombination reactions to go to completion. Proper positioning of dif in the chromosome and of FtsK at the septum is required to sense the multimeric state of newly replicated chromosomes and restrict complete Xer reactions to dimeric chromosomes.
Figures
References
-
- Arciszewska LK, Baker RA, Hallet B, Sherratt DJ. Coordinated control of XerC and XerD catalytic activities during Holliday junction resolution. J Mol Biol. 2000;299:391–403. - PubMed
-
- Austin S, Ziese M, Sternberg N. A novel role for site-specific recombination in maintenance of bacterial replicons. Cell. 1981;25:729–736. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases