

Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development
- PMID: 11114891
- PMCID: PMC317103
- DOI: 10.1101/gad.852200
Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development
Abstract
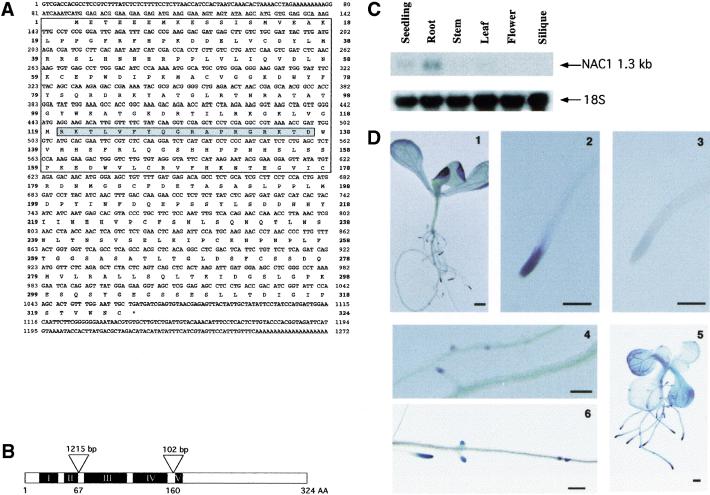
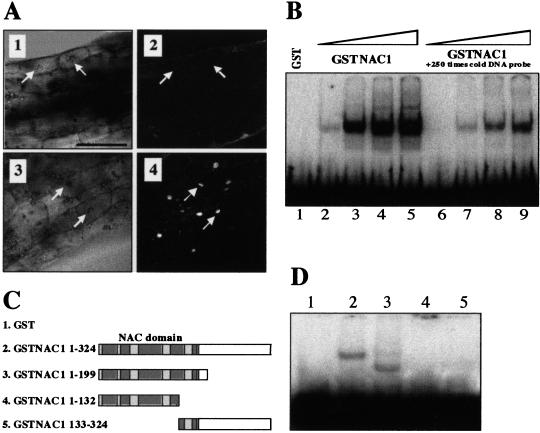
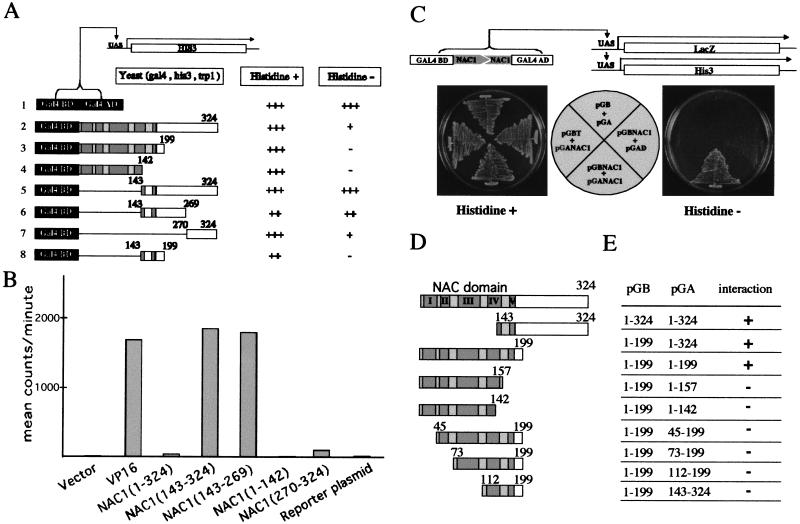
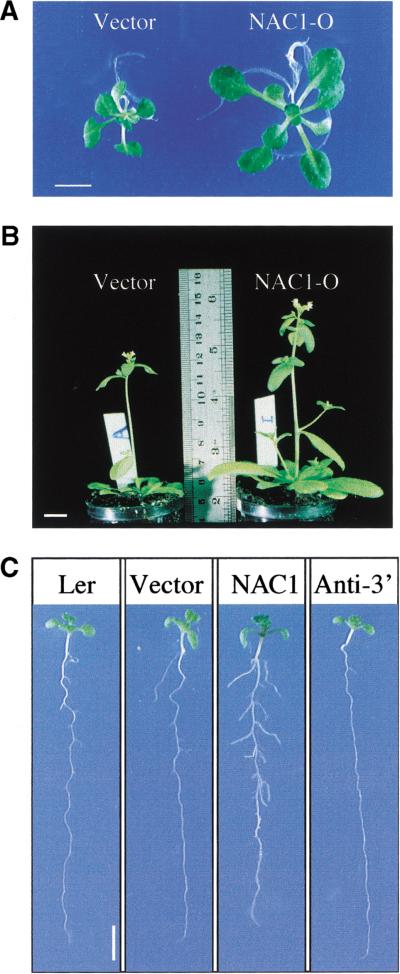
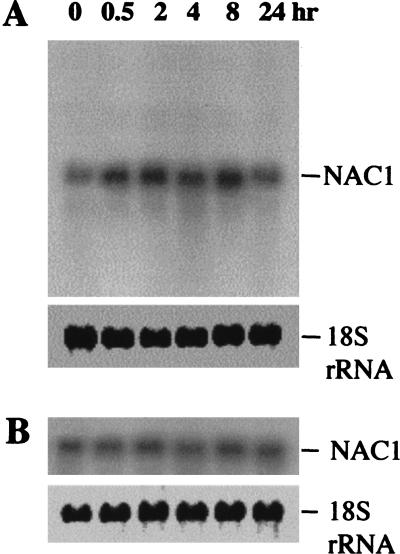
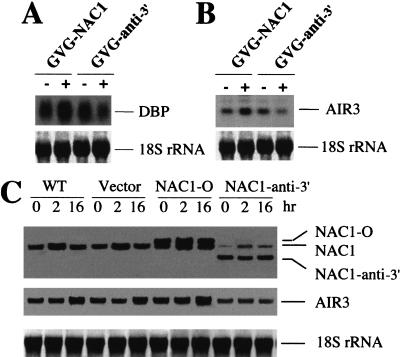
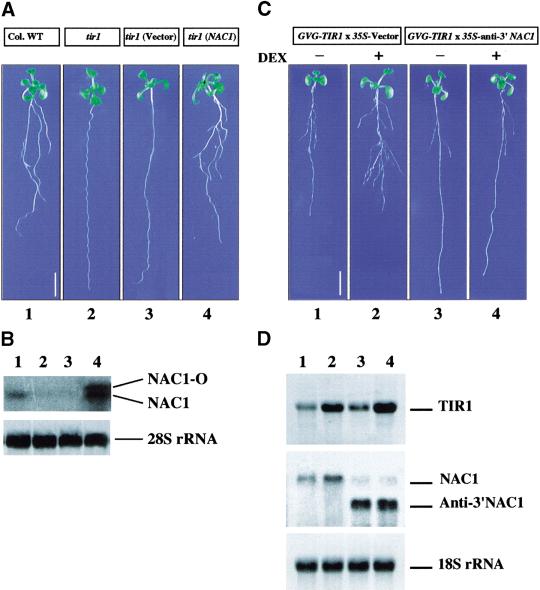
Auxin plays a key role in lateral root formation, but the signaling pathway for this process is poorly understood. We show here that NAC1, a new member of the NAC family, is induced by auxin and mediates auxin signaling to promote lateral root development. NAC1 is a transcription activator consisting of an N-terminal conserved NAC-domain that binds to DNA and a C-terminal activation domain. This factor activates the expression of two downstream auxin-responsive genes, DBP and AIR3. Transgenic plants expressing sense or antisense NAC1 cDNA show an increase or reduction of lateral roots, respectively. Finally, TIR1-induced lateral root development is blocked by expression of antisense NAC1 cDNA, and NAC1 overexpression can restore lateral root formation in the auxin-response mutant tir1, indicating that NAC1 acts downstream of TIR1.
Figures
References
-
- Aida M, Ishida T, Tasaka M. Shoot apical meristem and cotyledon formation during Arabidopsis embryogenesis: Interaction among the CUP-SHAPED COTYLEDON and SHOOT MERISTEMLESS genes. Development. 1999;126:1563–1570. - PubMed
-
- Aoyama T, Chua NH. A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 1997;11:605–612. - PubMed
-
- Bartel PL, Chien C-T, Sternglanz R, Fields S. Using the two-hybrid system to detect protein–protein interactions. In: In: Hartley DA, editor. Cellular interactions in development: A practical approach. Oxford: Oxford University Press; 1993a. pp. 153–179.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases