

The isolation and characterization in yeast of a gene for Arabidopsis S-adenosylmethionine:phospho-ethanolamine N-methyltransferase
- PMID: 11115895
- PMCID: PMC59876
- DOI: 10.1104/pp.124.4.1800
The isolation and characterization in yeast of a gene for Arabidopsis S-adenosylmethionine:phospho-ethanolamine N-methyltransferase
Abstract
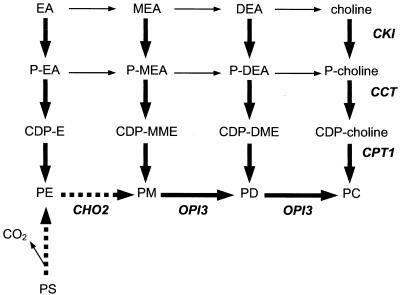
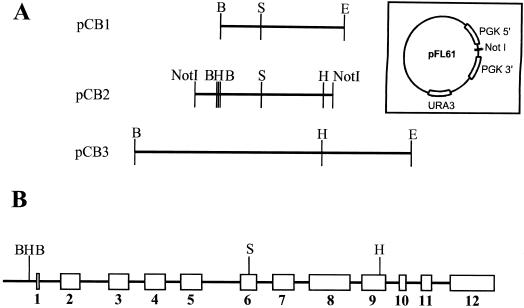
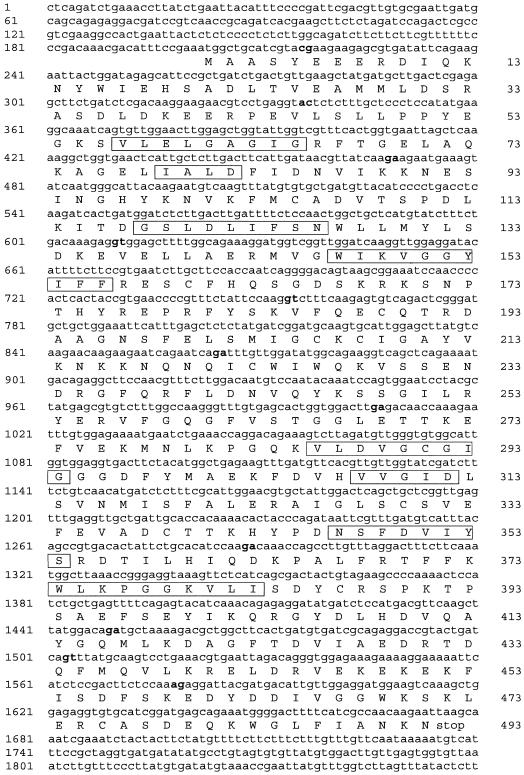
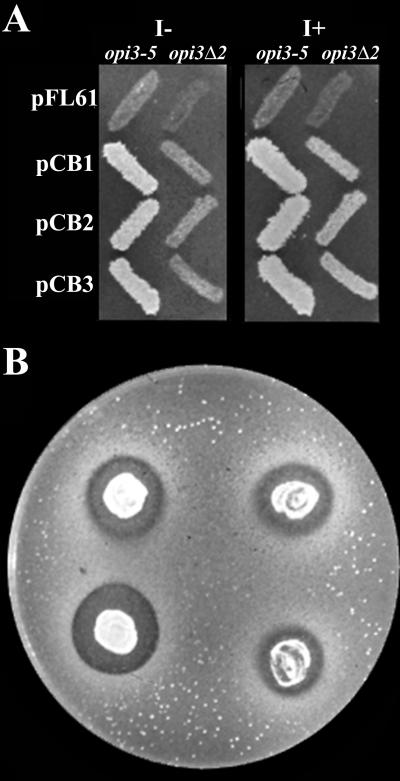
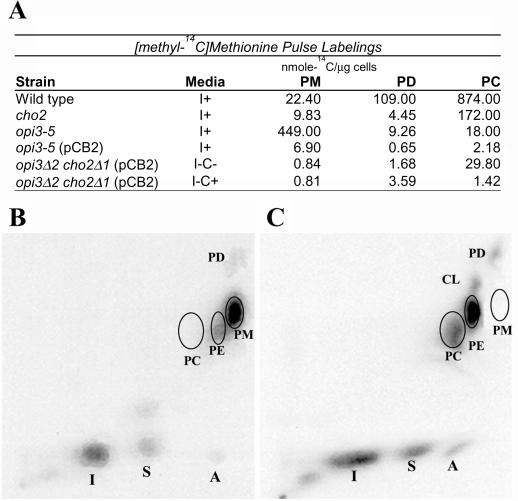
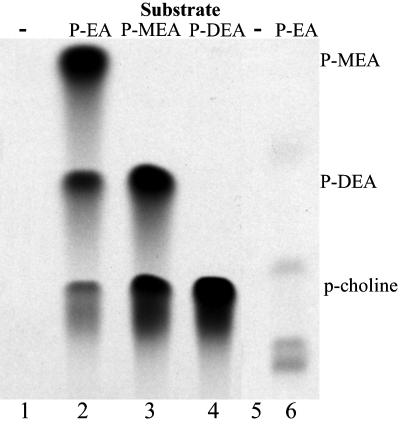
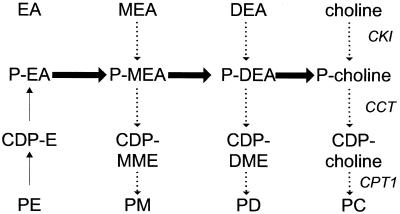
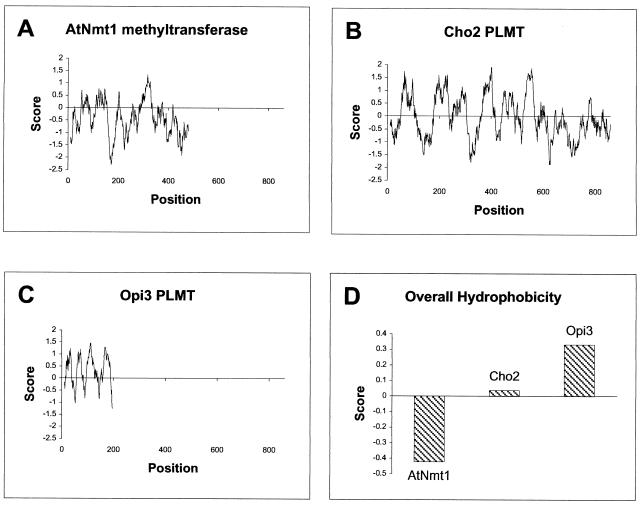
Saccharomyces cerevisiae opi3 mutant strains do not have the phospholipid N-methyltransferase that catalyzes the two terminal methylations in the phosphatidylcholine (PC) biosynthetic pathway. This results in a build up of the intermediate phosphatidylmonomethylethanolamine, causing a temperature-sensitive growth phenotype. An Arabidopsis cDNA library was used to isolate three overlapping plasmids that complemented the temperature-sensitive phenotype. Phospholipid analysis showed that the presence of the cloned cDNA caused a 65-fold reduction in the level of phosphatidylmonomethylethanolamine and a significant, though not equivalent, increase in the production of PC. Sequence analysis established that the cDNA was not homologous to OPI3 or to CHO2, the only other yeast phospholipid N-methyltransferase, but was similar to several other classes of methyltransferases. S-adenosyl-Met:phospho-base N-methyltransferase assays revealed that the cDNA catalyzed the three sequential methylations of phospho-ethanolamine to form phospho-choline. Phospho-choline is converted to PC by the CDP-choline pathway, explaining the phenotype conferred upon the yeast mutant strain by the cDNA. In accordance with this the gene has been named AtNMT1. The identification of this enzyme and the failure to isolate a plant phospholipid N-methyltransferase suggests that there are fundamental differences between the pathways utilized by yeast and by some plants for synthesis of PC.
Figures

References
-
- Bjellqvist B, Basse B, Olsen E, Celis JE. Reference points for comparisons of two-dimensional maps of proteins from different human cell types defined in a pH scale where isoelectric points correlate with polypeptide compositions. Electrophoresis. 1994;15:529–539. - PubMed
-
- Bjellqvist B, Hughes GJ, Pasquali C, Paquet N, Ravier F, Sanchez J, Frutiger S, Hochstrasser DF. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis. 1993;14:1023–1031. - PubMed
-
- Boeke JD, LaCroute F, Fink GR. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
