

Phenotypic analysis and virulence of Candida albicans LIG4 mutants
- PMID: 11119499
- PMCID: PMC97865
- DOI: 10.1128/IAI.69.01.137-147.2001
Phenotypic analysis and virulence of Candida albicans LIG4 mutants
Abstract
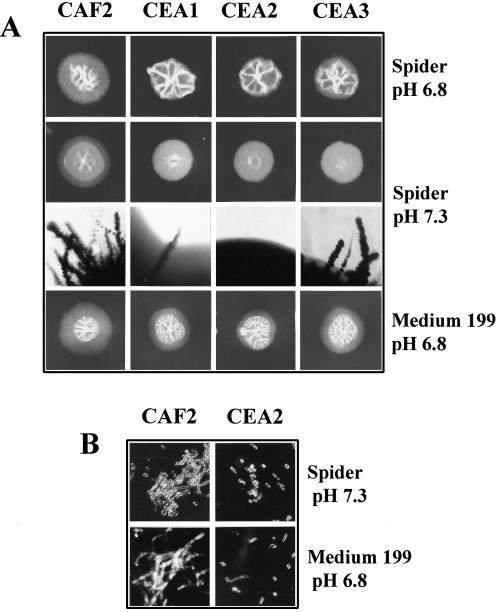
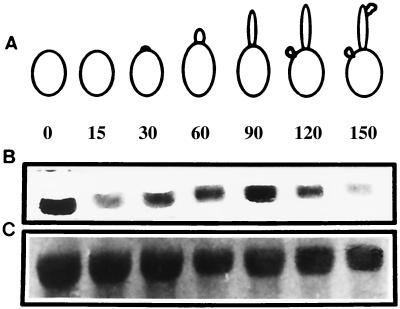
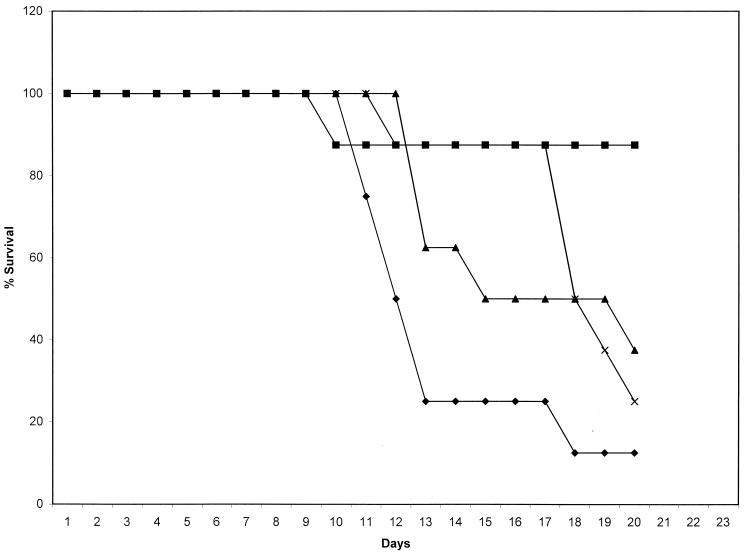
In previous studies, we reported the isolation and preliminary characterization of a DNA ligase-encoding gene of Candida albicans. This gene (LIG4) is the structural and functional homologue of both yeast and human ligase IV, which is involved in nonhomologous end joining (NHEJ) of DNA double-strand breaks. In the present study, we have shown that there are no other LIG4 homologues in C. albicans. In order to study the function of LIG4 in morphogenesis and virulence, we constructed gene deletions. LIG4 transcript levels were reduced in the heterozygote and were completely absent in null strains. Concomitantly, the heterozygote showed a pronounced defect in myceliation, which was slightly greater in the null strain. This was true with several solid and liquid media, such as Spider medium, medium 199, and 2% glucose-1% yeast extract-2% Bacto Peptone, at several pHs. Reintroduction of the wild-type allele into the null mutant partially restored the ability of cells to form hyphae. In agreement with the positive role of LIG4 in morphogenesis, we detected a significant rise in mRNA levels during the morphological transition. LIG4 is not essential for DNA replication or for the repair of DNA damage induced by ionizing radiation or UV light, indicating that these lesions are repaired primarily by homologous recombination. However, our data show that the NHEJ apparatus of C. albicans may control morphogenesis in this diploid organism. In addition, deletion of one or both copies of LIG4 resulted in attenuation of virulence in a murine model of candidiasis.
Figures


References
-
- Andaluz E, Larriba G, Calderone R. A Candida albicans gene encoding a DNA ligase. Yeast. 1996;12:893–898. - PubMed
-
- Andaluz E, Ciudad A, Rubio Coque J, Calderone R, Larriba G. Cell cycle regulation of a DNA ligase-encoding gene (CaLIG4) from Candida albicans. Yeast. 1999;15:1199–1210. - PubMed
-
- Boeke J D, LaCroute F, Fink G R. A positive selection for mutants lacking orotidine-5-phosphate carboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet. 1984;197:345–346. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
