

DNA condensation in two dimensions
- PMID: 11121015
- PMCID: PMC18867
- DOI: 10.1073/pnas.97.26.14046
DNA condensation in two dimensions
Abstract
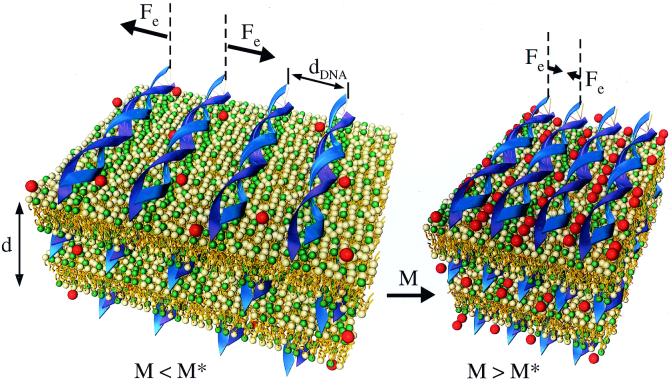
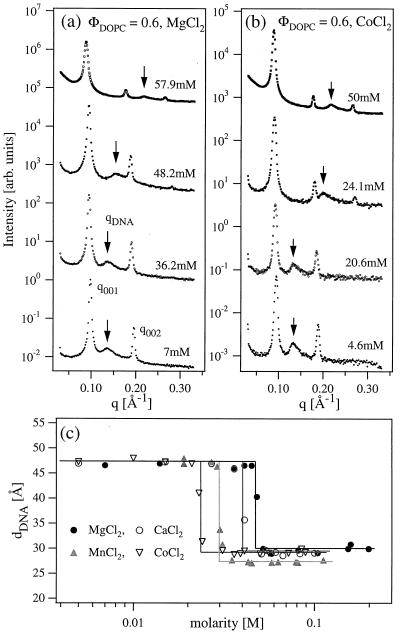
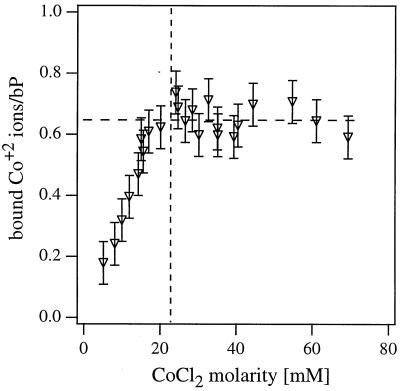
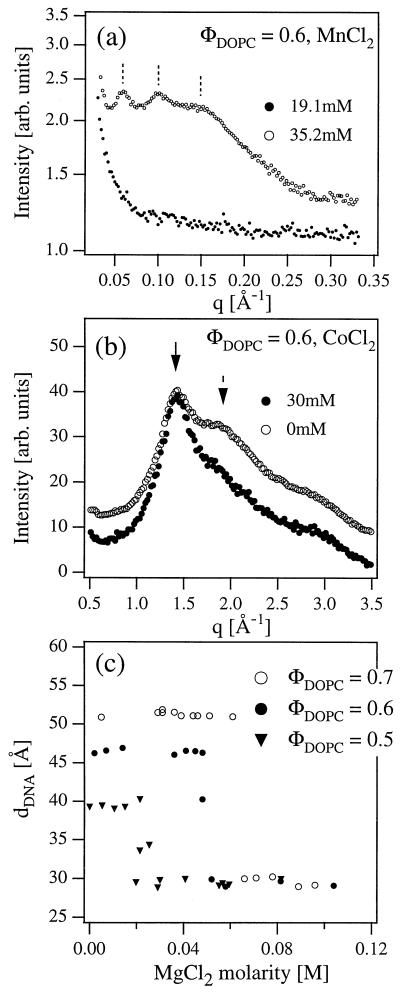
We have found that divalent electrolyte counterions common in biological cells (Ca(2+), Mg(2+), and Mn(2+) ) can condense anionic DNA molecules confined to two-dimensional cationic surfaces. DNA-condensing agents in vivo include cationic histones and polyamines spermidine and spermine with sufficiently high valence (Z) 3 or larger. In vitro studies show that electrostatic forces between DNA chains in bulk aqueous solution containing divalent counterions remain purely repulsive, and DNA condensation requires counterion valence Z >/= 3. In striking contrast to bulk behavior, synchrotron x-ray diffraction and optical absorption experiments show that above a critical divalent counterion concentration the electrostatic forces between DNA chains adsorbed on surfaces of cationic membranes reverse from repulsive to attractive and lead to a chain collapse transition into a condensed phase of DNA tethered by divalent counterions. This demonstrates the importance of spatial dimensionality to intermolecular interactions where nonspecific counterion-induced electrostatic attractions between the like-charged polyelectrolytes overwhelm the electrostatic repulsions on a surface for Z = 2. This new phase, with a one-dimensional counterion liquid trapped between DNA chains at a density of 0.63 counterions per DNA bp, represents the most compact state of DNA on a surface in vitro and suggests applications in high-density storage of genetic information and organo-metallic materials processing.
Figures
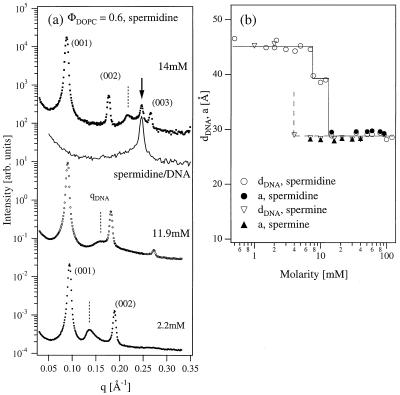
Similar articles
-
Thermodynamics of cation-induced DNA condensation.J Biomol Struct Dyn. 1991 Aug;9(1):159-67. doi: 10.1080/07391102.1991.10507900. J Biomol Struct Dyn. 1991. PMID: 1838258
-
A close encounter with DNA.Eur Biophys J. 2003 Sep;32(6):585-7. doi: 10.1007/s00249-003-0307-x. Epub 2003 Aug 8. Eur Biophys J. 2003. PMID: 12908089
-
Roles of anionic and cationic template components in biomineralization of CdS nanorods using self-assembled DNA-membrane complexes.J Am Chem Soc. 2004 Nov 3;126(43):14157-65. doi: 10.1021/ja046718m. J Am Chem Soc. 2004. PMID: 15506781
-
DNA condensation by multivalent cations.Biopolymers. 1997;44(3):269-82. doi: 10.1002/(SICI)1097-0282(1997)44:3<269::AID-BIP6>3.0.CO;2-T. Biopolymers. 1997. PMID: 9591479 Review.
-
The contribution of transient counterion imbalances to DNA bending fluctuations.Biophys J. 2006 May 1;90(9):3208-15. doi: 10.1529/biophysj.105.078865. Epub 2006 Feb 3. Biophys J. 2006. PMID: 16461401 Free PMC article. Review.
Cited by
-
A novel polymer-lipid hybrid nanoparticle for efficient nonviral gene delivery.Acta Pharmacol Sin. 2010 Apr;31(4):509-14. doi: 10.1038/aps.2010.15. Epub 2010 Mar 29. Acta Pharmacol Sin. 2010. PMID: 20348944 Free PMC article.
-
Divalent counterion-induced condensation of triple-strand DNA.Proc Natl Acad Sci U S A. 2010 Dec 14;107(50):21482-6. doi: 10.1073/pnas.1003374107. Epub 2010 Nov 22. Proc Natl Acad Sci U S A. 2010. PMID: 21098260 Free PMC article.
-
Ion-mediated nucleic acid helix-helix interactions.Biophys J. 2006 Jul 15;91(2):518-36. doi: 10.1529/biophysj.106.084285. Epub 2006 Apr 28. Biophys J. 2006. PMID: 16648172 Free PMC article.
-
Polynucleotide adsorption to negatively charged surfaces in divalent salt solutions.Biophys J. 2006 Feb 15;90(4):1164-74. doi: 10.1529/biophysj.105.070649. Biophys J. 2006. PMID: 16449197 Free PMC article.
-
A review of heavy metal cation binding to deoxyribonucleic acids for the creation of chemical sensors.Biophys Rev. 2018 Oct;10(5):1401-1414. doi: 10.1007/s12551-018-0455-y. Epub 2018 Sep 18. Biophys Rev. 2018. PMID: 30229467 Free PMC article. Review.
References
-
- Lewin B. Genes VII. London: Oxford Univ. Press; 1999.
-
- Bloomfield V A. Biopolymers. 1997;44:269–282. - PubMed
-
- Wilson R W, Bloomfield V A. Biochemistry. 1979;18:2192–2196. - PubMed
-
- Bloomfield V A. Curr Opin Struct Biol. 1996;6:334–343. - PubMed
-
- Pelta J, Livolant F, Sikorav J-L. J Biol Chem. 1996;271:5656–5666. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
