

Controlling gene expression in yeast by inducible site-specific recombination
- PMID: 11121495
- PMCID: PMC115252
- DOI: 10.1093/nar/28.24.e108
Controlling gene expression in yeast by inducible site-specific recombination
Abstract
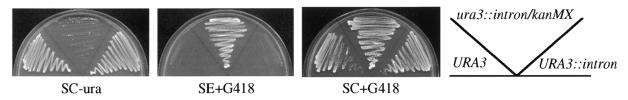
An intron module was developed for Saccharomyces cerevisiae that imparts conditional gene regulation. The kanMX marker, flanked by loxP sites for the Cre recombinase, was embedded within the ACT1 intron and the resulting module was targeted to specific genes by PCR-mediated gene disruption. Initially, recipient genes were inactivated because the loxP-kanMX-loxP cassette prevented formation of mature transcripts. However, expression was restored by Cre-mediated site-specific recombination, which excised the loxP-kanMX-loxP cassette to generate a functional intron that contained a single loxP site. Cre recombinase activity was controlled at the transcriptional level by a GAL1::CRE expression vector or at the enzymatic level by fusing the protein to the hormone-dependent regulatory domain of the estrogen receptor. Negative selection against leaky pre-excision events was achieved by growing cells in modified minimal media that contained geneticin (G418). Advantages of this gene regulation technique, which we term the conditional knock-out approach, are that (i) modified genes are completely inactivated prior to induction, (ii) modified genes are induced rapidly to expression levels that compare to their unmodified counterparts, and (iii) it is easy to use and generally applicable.
Figures




References
-
- Mumberg D., Müller,R. and Funk,M. (1995) Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds. Gene, 156, 119–122. - PubMed
-
- Rönicke V., Graulich,W., Mumberg,D., Müller,R. and Funk,M. (1997) Use of conditional promoters for expression of heterologous proteins in Saccharomyces cerevisiae. Methods Enzymol., 283, 313–322. - PubMed
-
- Labbé S. and Thiele,D.J. (1999) Copper ion inducible and repressible promoter systems in yeast. Methods Enzymol., 306, 145–153. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
