

Wild-type huntingtin reduces the cellular toxicity of mutant huntingtin in vivo
- PMID: 11133364
- PMCID: PMC1235265
- DOI: 10.1086/318207
Wild-type huntingtin reduces the cellular toxicity of mutant huntingtin in vivo
Abstract
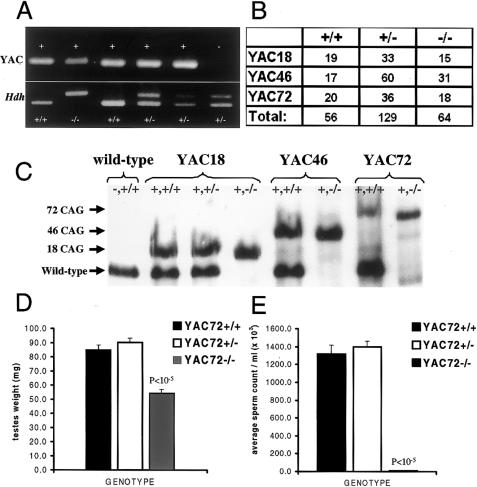
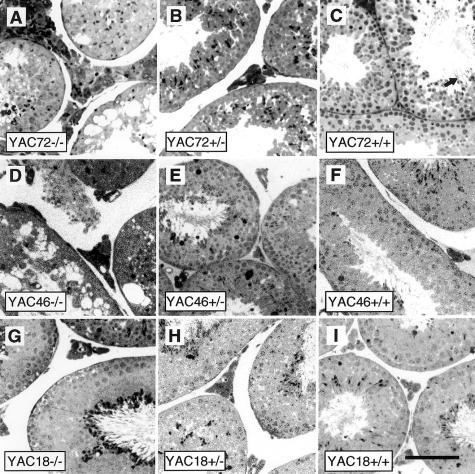
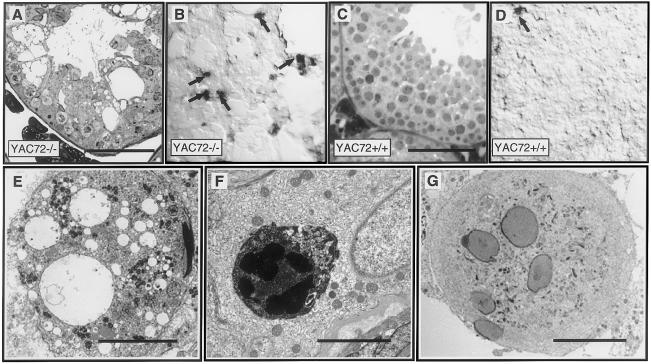
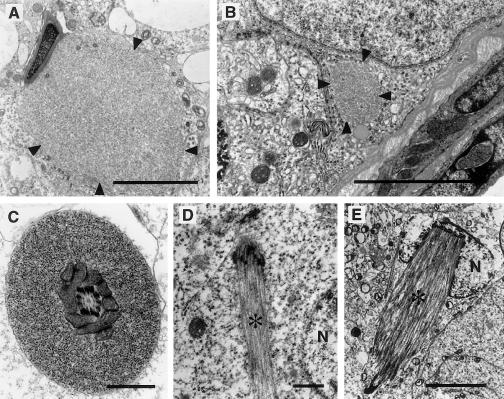
We have developed yeast artificial chromosome (YAC) transgenic mice expressing normal (YAC18) and mutant (YAC46 or YAC72) human huntingtin (htt), in a developmental- and tissue-specific manner, that is identical to endogenous htt. YAC72 mice develop selective degeneration of medium spiny projection neurons in the lateral striatum, similar to what is observed in Huntington disease. Mutant human htt expressed by YAC transgenes can compensate for the absence of endogenous htt and can rescue the embryonic lethality that characterizes mice homozygous for targeted disruption of the endogenous Hdh gene (-/-). YAC72 mice lacking endogenous htt (YAC72 -/-) manifest a novel phenotype characterized by infertility, testicular atrophy, aspermia, and massive apoptotic cell death in the testes. The testicular cell death in YAC72 -/- mice can be markedly reduced by increasing endogenous htt levels. YAC72 mice with equivalent levels of both wild-type and mutant htt (YAC72 +/+) breed normally and have no evidence of increased testicular cell death. Similar findings are seen in YAC46 -/- mice compared with YAC46 +/+ mice, in which wild-type htt can completely counteract the proapoptotic effects of mutant htt. YAC18 -/- mice display no evidence of increased cellular apoptosis, even in the complete absence of endogenous htt, demonstrating that the massive cellular apoptosis observed in YAC46 -/- mice and YAC72 -/- mice is polyglutamine-mediated toxicity from the mutant transgene. These data provide the first direct in vivo evidence of a role for wild-type htt in decreasing the cellular toxicity of mutant htt.
Figures

References
Electronic-Database Information
-
- Genome Database, The, http://www.gdb.org (for HD [accession number 119307])
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for HD [MIM 143100])
References
-
- Andrew SE, Goldberg YP, Kremer B, Telenius H, Theilmann J, Adam S, Starr E, Squitieri F, Lin B, Kalchman MA, Graham RK, Hayden MR (1993) The relationship between trinucleotide (CAG) repeat length and clinical features of Huntington's disease. Nat Genet 4:398–403 - PubMed
-
- Becher MW, Kotzuk JA, Sharp AH, Davies SW, Bates GP, Price DL, Ross CR (1998) Intranuclear neuronal inclusions in Huntington`s disease and dentatorubral and pallidoluysian atrophy: correlation between density of inclusions and IT15 triplet repeat length. Neurobiol Dis 4:387–397 - PubMed
-
- Burke JR, Enghild JJ, Martin ME, Jou Y-S, Myers RM, Roses AD, Vance JM, Strittmatter WJ (1996) Huntingtin and DRPLA proteins selectively interact with the enzyme GADPH. Nat Med 2:347–349 - PubMed
-
- Cooper JK, Schilling G, Peters MF, Herring WJ, Sharp AH, Kaminsky Z, Masone J, Khan FA, Delanoy M, Borschelt DR, Dawson VL, Dawson TM, Ross CA (1998) Truncated N-terminal fragments of huntingtin with expanded glutamine repeats form nuclear and cytoplasmic aggregates in cell culture. Hum Mol Genet 7:783–790 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
