

Light adaptation in Drosophila photoreceptors: II. Rising temperature increases the bandwidth of reliable signaling
- PMID: 11134229
- PMCID: PMC2232470
- DOI: 10.1085/jgp.117.1.27
Light adaptation in Drosophila photoreceptors: II. Rising temperature increases the bandwidth of reliable signaling
Abstract
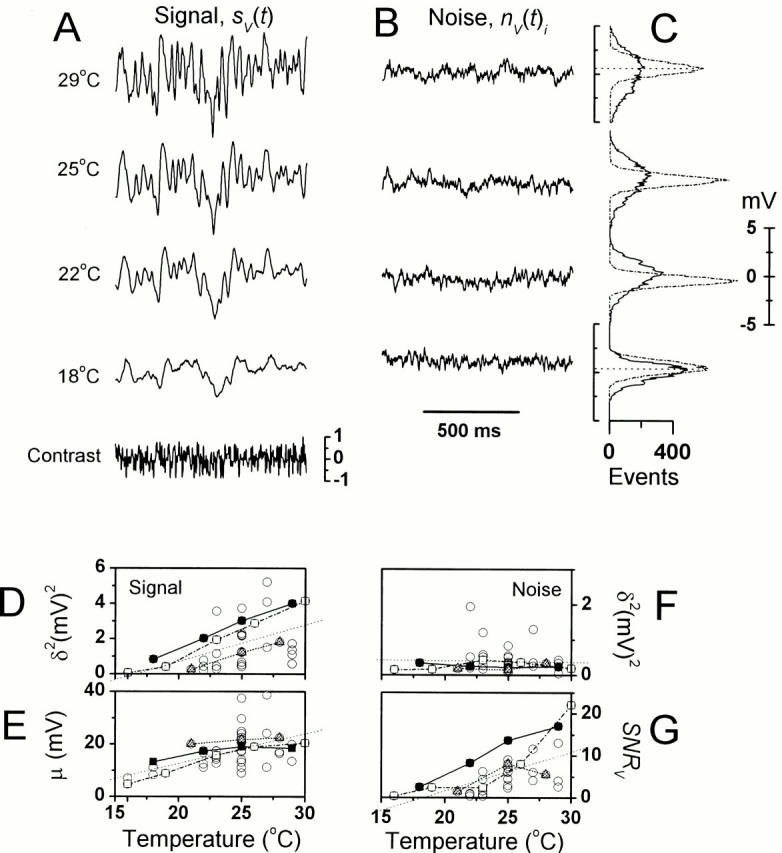
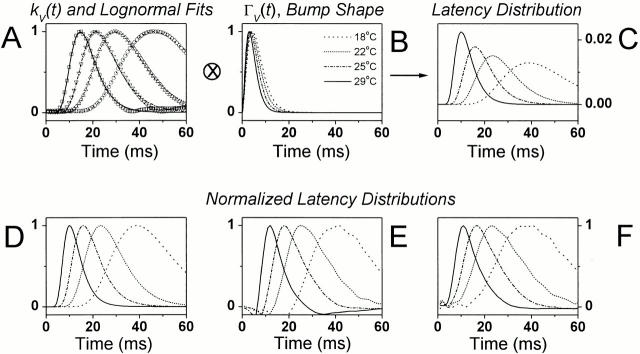
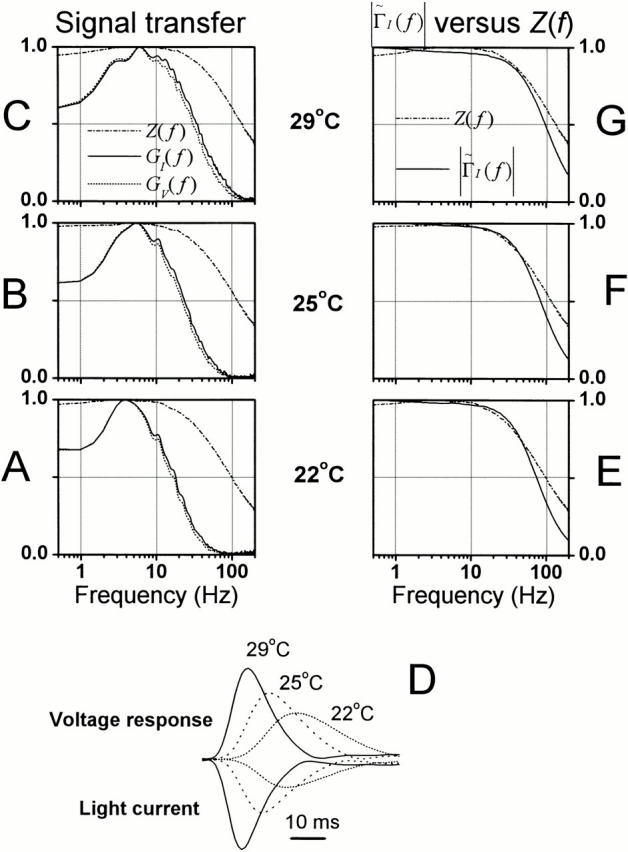
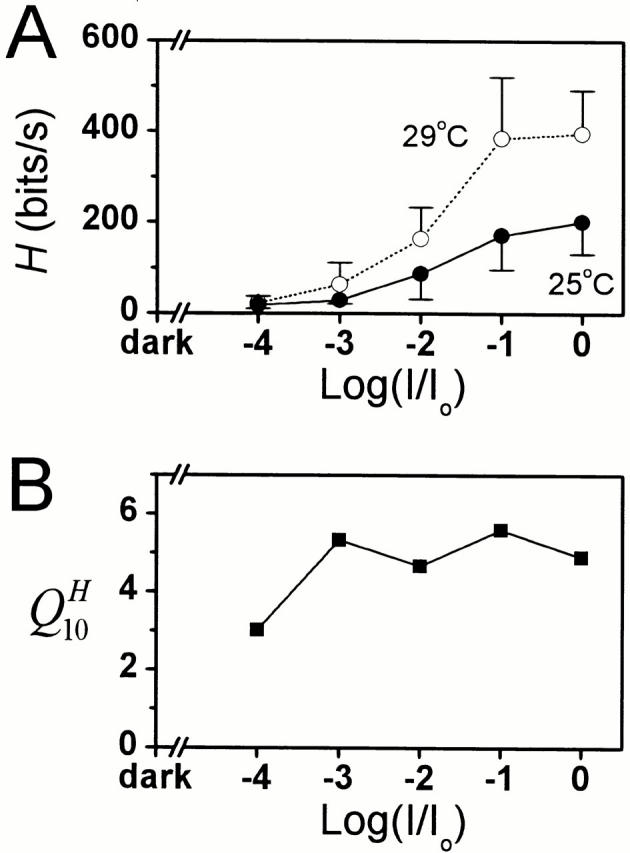
It is known that an increase in both the mean light intensity and temperature can speed up photoreceptor signals, but it is not known whether a simultaneous increase of these physical factors enhances information capacity or leads to coding errors. We studied the voltage responses of light-adapted Drosophila photoreceptors in vivo from 15 to 30 degrees C, and found that an increase in temperature accelerated both the phototransduction cascade and photoreceptor membrane dynamics, broadening the bandwidth of reliable signaling with an effective Q(10) for information capacity of 6.5. The increased fidelity and reliability of the voltage responses was a result of four factors: (1) an increased rate of elementary response, i.e., quantum bump production; (2) a temperature-dependent acceleration of the early phototransduction reactions causing a quicker and narrower dispersion of bump latencies; (3) a relatively temperature-insensitive light-adapted bump waveform; and (4) a decrease in the time constant of the light-adapted photoreceptor membrane, whose filtering matched the dynamic properties of the phototransduction noise. Because faster neural processing allows faster behavioral responses, this improved performance of Drosophila photoreceptors suggests that a suitably high body temperature offers significant advantages in visual performance.
Figures






Similar articles
-
Light adaptation in Drosophila photoreceptors: I. Response dynamics and signaling efficiency at 25 degrees C.J Gen Physiol. 2001 Jan;117(1):3-25. doi: 10.1085/jgp.117.1.3. J Gen Physiol. 2001. PMID: 11134228 Free PMC article.
-
Latency of phototransduction limits transfer of higher-frequency signals in cockroach photoreceptors.J Neurophysiol. 2020 Jan 1;123(1):120-133. doi: 10.1152/jn.00365.2019. Epub 2019 Nov 13. J Neurophysiol. 2020. PMID: 31721631
-
Visual coding in locust photoreceptors.PLoS One. 2008 May 14;3(5):e2173. doi: 10.1371/journal.pone.0002173. PLoS One. 2008. PMID: 18478123 Free PMC article.
-
How a fly photoreceptor samples light information in time.J Physiol. 2017 Aug 15;595(16):5427-5437. doi: 10.1113/JP273645. Epub 2017 Apr 17. J Physiol. 2017. PMID: 28233315 Free PMC article. Review.
-
Phototransduction in Drosophila.Sci China Life Sci. 2012 Jan;55(1):27-34. doi: 10.1007/s11427-012-4272-4. Epub 2012 Feb 8. Sci China Life Sci. 2012. PMID: 22314488 Review.
Cited by
-
Network adaptation improves temporal representation of naturalistic stimuli in Drosophila eye: I dynamics.PLoS One. 2009;4(1):e4307. doi: 10.1371/journal.pone.0004307. Epub 2009 Jan 30. PLoS One. 2009. PMID: 19180196 Free PMC article.
-
The effects of temperature on signalling in ocellar neurons of the desert locust, Schistocerca gregaria.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011 Nov;197(11):1083-96. doi: 10.1007/s00359-011-0669-y. Epub 2011 Aug 10. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2011. PMID: 21830121
-
Microsaccadic sampling of moving image information provides Drosophila hyperacute vision.Elife. 2017 Sep 5;6:e26117. doi: 10.7554/eLife.26117. Elife. 2017. PMID: 28870284 Free PMC article.
-
Parallel Gene Expression Differences between Low and High Latitude Populations of Drosophila melanogaster and D. simulans.PLoS Genet. 2015 May 7;11(5):e1005184. doi: 10.1371/journal.pgen.1005184. eCollection 2015 May. PLoS Genet. 2015. PMID: 25950438 Free PMC article.
-
Predicting individual neuron responses with anatomically constrained task optimization.Curr Biol. 2021 Sep 27;31(18):4062-4075.e4. doi: 10.1016/j.cub.2021.06.090. Epub 2021 Jul 28. Curr Biol. 2021. PMID: 34324832 Free PMC article.
References
-
- Aho A.C., Donner K., Hyden C., Larsen L.O., Reuter T. Low retinal noise in animals with low body-temperature allows high visual sensitivity. Nature. 1988;334:348–350. - PubMed
-
- French A.S. Phototransduction in the fly compound eye exhibits temporal resonances and a pure time delay. Nature. 1980;283:200–202. - PubMed
-
- Heinrich B. The hod-blooded insects 1993. Springer-Verlag; Berlin: pp. 600 pp
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
