

Interleukin-6-induced STAT3 and AP-1 amplify hepatocyte nuclear factor 1-mediated transactivation of hepatic genes, an adaptive response to liver injury
- PMID: 11134330
- PMCID: PMC86585
- DOI: 10.1128/MCB.21.2.414-424.2001
Interleukin-6-induced STAT3 and AP-1 amplify hepatocyte nuclear factor 1-mediated transactivation of hepatic genes, an adaptive response to liver injury
Abstract
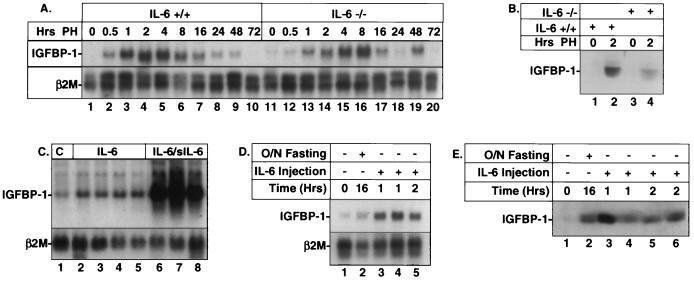
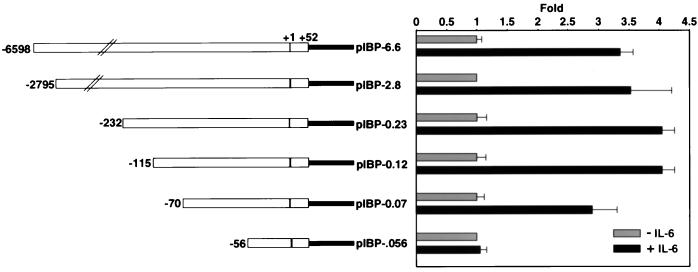
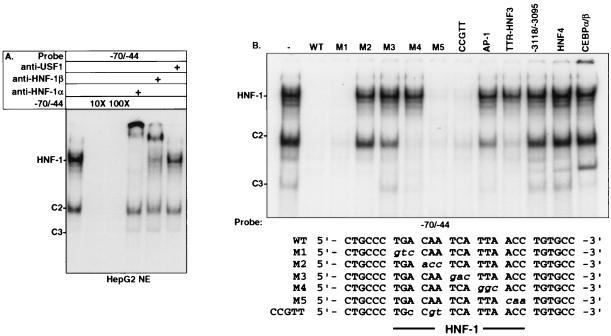
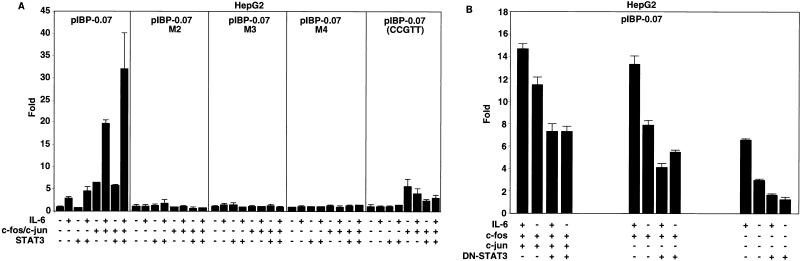
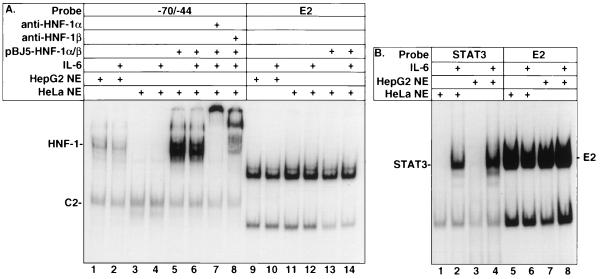
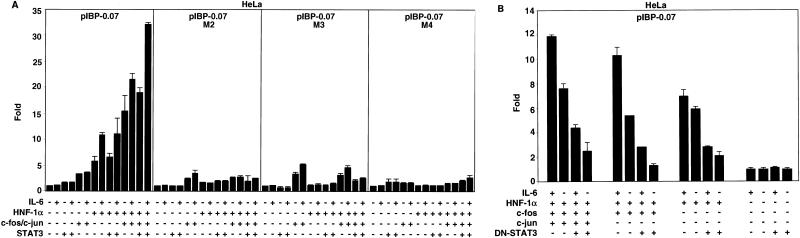
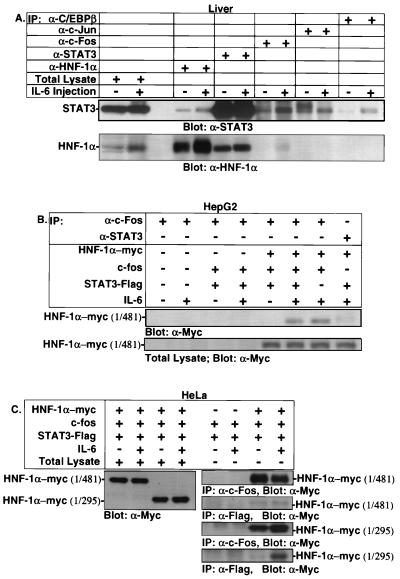
Following hepatic injury or stress, gluconeogenic and acute-phase response genes are rapidly upregulated to restore metabolic homeostasis and limit tissue damage. Regulation of the liver-restricted insulin-like growth factor binding protein 1 (IGFBP-1) gene is dramatically altered by changes in the metabolic state and hepatectomy, and thus it provided an appropriate reporter to assess the transcriptional milieu in the liver during repair and regeneration. The cytokine interleukin-6 (IL-6) is required for liver regeneration and repair, and it transcriptionally upregulates a vast array of genes during liver growth by unknown mechanisms. Evidence for a biologic role of IL-6 in IGFBP-1 upregulation was demonstrated by increased expression of hepatic IGFBP-1 in IL-6 transgenic and following injection of IL-6 into nonfasting animals and its reduced expression in IL-6(-/-) livers posthepatectomy. In both hepatic and nonhepatic cells, IL-6 -mediated IGFBP-1 promoter activation was via an intact hepatocyte nuclear factor 1 (HNF-1) site and was dependent on the presence of endogenous liver factor HNF-1 and induced factors STAT3 and AP-1 (c-Fos/c-Jun). IL-6 acted through the STAT3 pathway, as dominant negative STAT3 completely blocked IL-6-mediated stimulation of the IGFBP-1 promoter via the HNF-1 site. HNF-1/c-Fos and HNF-1/STAT3 protein complexes were detected in mouse livers and in hepatic and nonhepatic cell lines overexpressing STAT3/c-Fos/HNF-1. Similar regulation was demonstrated using glucose-6-phosphatase and alpha-fibrinogen promoters, indicating that HNF-1/IL-6/STAT3/AP-1-mediated transactivation of hepatic gene expression is a general phenomenon after liver injury. These results demonstrate that the two classes of transcription factors, growth induced (STAT3 and AP-1) and tissue specific (HNF-1), can interact as an adaptive response to liver injury to amplify expression of hepatic genes important for the homeostatic response during organ repair.
Figures
References
-
- Babajko S, Groyer A. Interplay of liver-enriched trans-acting factors, DBP and HNF1 in the transactivation of human IGFBP-1 promoter. Biochem Biophys Res Commun. 1993;196:480–486. - PubMed
-
- Cereghini S. Liver-enriched transcription factors and hepatocyte differentiation. FASEB J. 1996;10:267–282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous