

Nucleocytoplasmic distribution of budding yeast protein kinase A regulatory subunit Bcy1 requires Zds1 and is regulated by Yak1-dependent phosphorylation of its targeting domain
- PMID: 11134339
- PMCID: PMC86612
- DOI: 10.1128/MCB.21.2.511-523.2001
Nucleocytoplasmic distribution of budding yeast protein kinase A regulatory subunit Bcy1 requires Zds1 and is regulated by Yak1-dependent phosphorylation of its targeting domain
Abstract
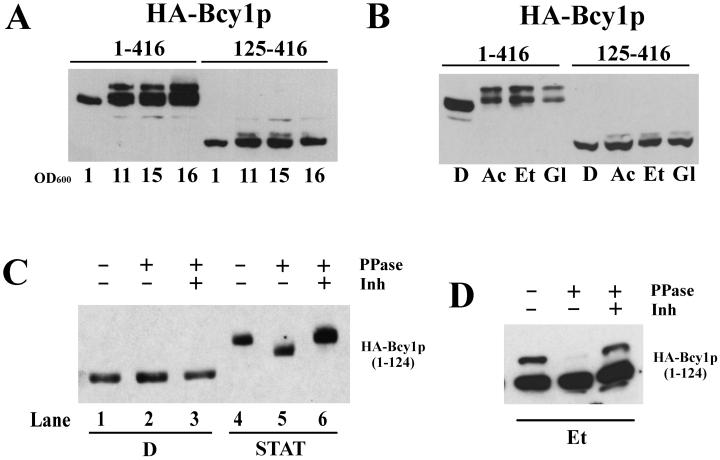
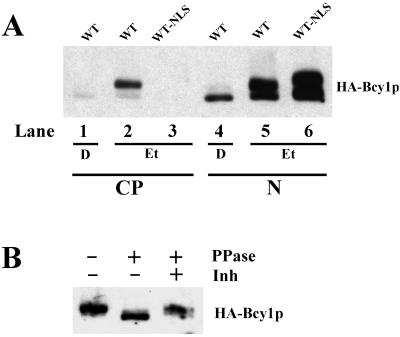
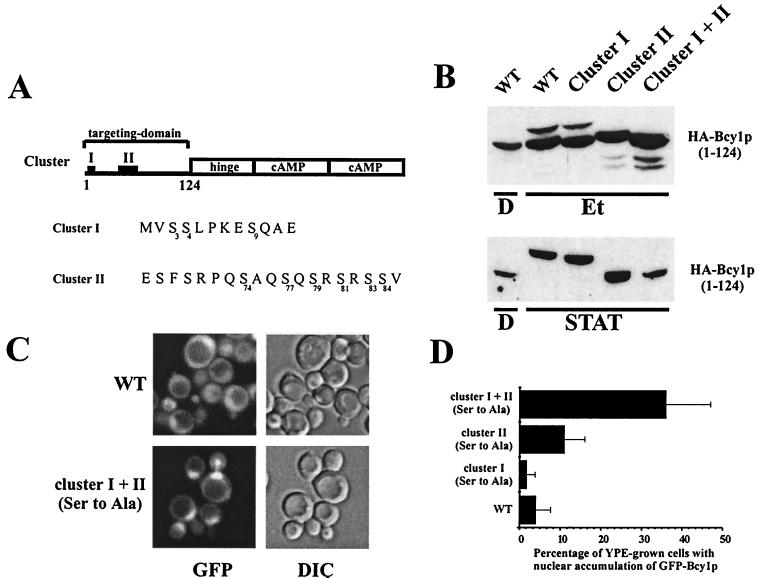
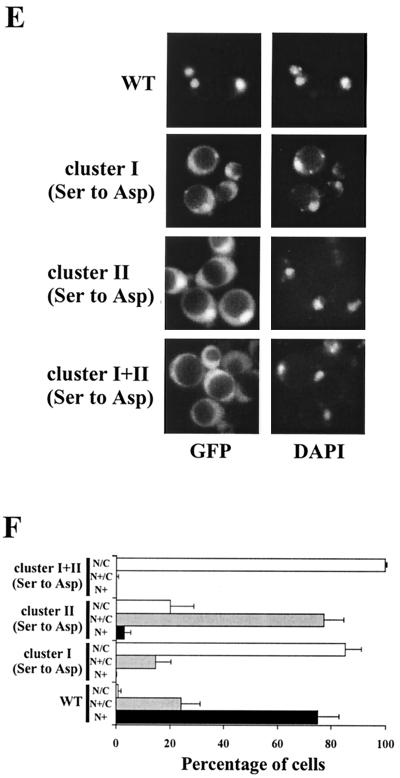
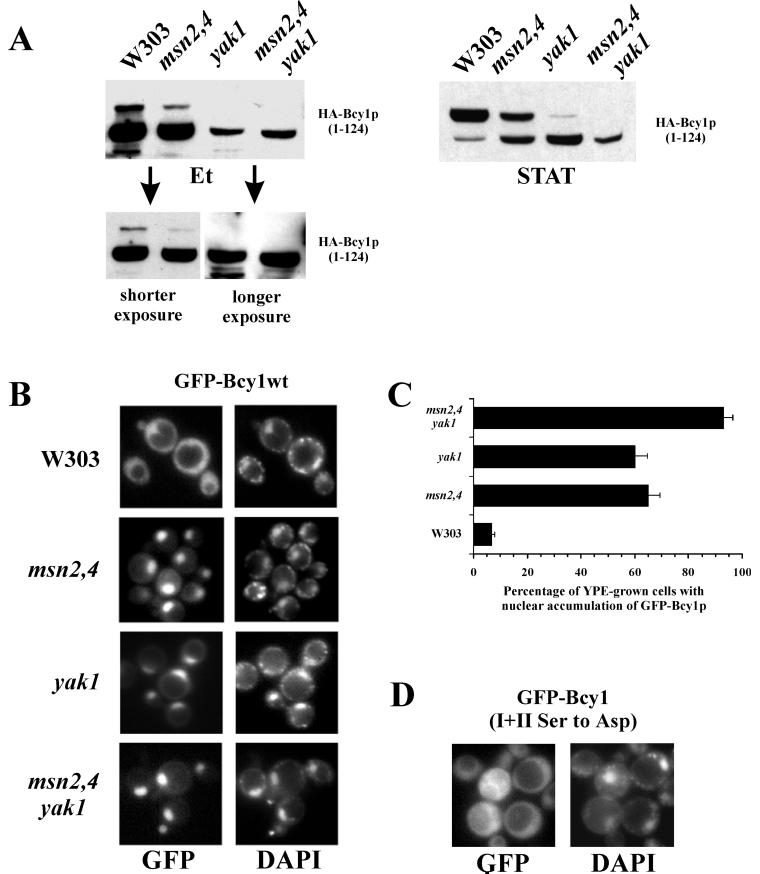
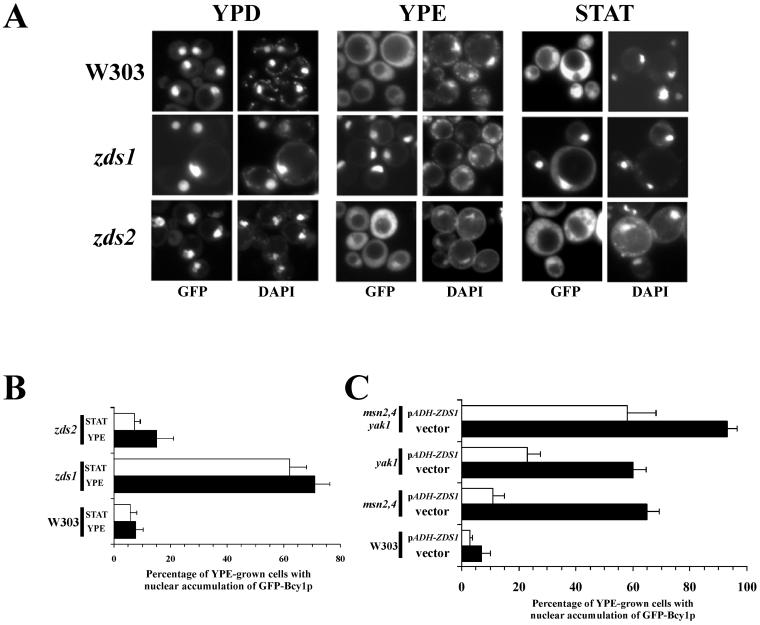
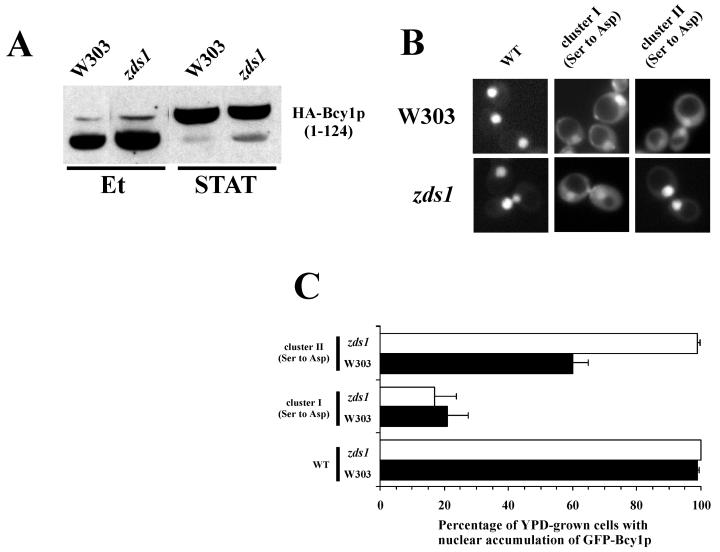
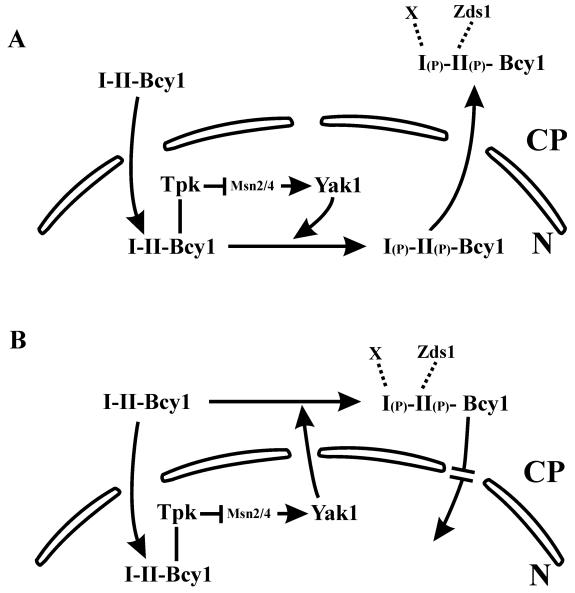
In Saccharomyces cerevisiae the subcellular distribution of Bcy1 is carbon source dependent. In glucose-grown cells, Bcy1 is almost exclusively nuclear, while it appears more evenly distributed between nucleus and cytoplasm in carbon source-derepressed cells. Here we show that phosphorylation of its N-terminal domain directs Bcy1 to the cytoplasm. Biochemical fractionation revealed that the cytoplasmic fraction contains mostly phosphorylated Bcy1, whereas unmodified Bcy1 is predominantly present in the nuclear fraction. Site-directed mutagenesis of two clusters (I and II) of serines near the N terminus to alanine resulted in an enhanced nuclear accumulation of Bcy1 in ethanol-grown cells. In contrast, substitutions to Asp led to a dramatic increase of cytoplasmic localization in glucose-grown cells. Bcy1 modification was found to be dependent on Yak1 kinase and, consequently, in ethanol-grown yak1 cells the Bcy1 remained nuclear. A two-hybrid screen aimed to isolate genes encoding proteins that interact with the Bcy1 N-terminal domain identified Zds1. In ethanol-grown zds1 cells, cytoplasmic localization of Bcy1 was largely absent, while overexpression of ZDS1 led to increased cytoplasmic Bcy1 localization. Zds1 does not regulate Bcy1 modification since this was found to be unaffected in zds1 cells. However, in zds1 cells cluster II-mediated, but not cluster I-mediated, cytoplasmic localization of Bcy1 was found to be absent. Altogether, these results suggest that Zds1-mediated cytoplasmic localization of Bcy1 is regulated by carbon source-dependent phosphorylation of cluster II serines, while cluster I acts in a Zds1-independent manner.
Figures
References
-
- Ausubel F M, Brent B, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: Green Publishing Associates and Wiley Interscience; 1993.
-
- Beebe S J. The cAMP-dependent protein kinases and cAMP signal transduction. Semin Cancer Biol. 1994;5:285–294. - PubMed
-
- Bregman D B, Bhattacharyya N, Rubin C S. High affinity binding protein for the regulatory subunit of cAMP-dependent protein kinase II-B. Cloning, characterization, and expression of cDNAs for rat brain P150. J Biol Chem. 1989;264:4648–4656. - PubMed
-
- Colledge M, Scott J D. AKAPs: from structure to function. Trends Cell Biol. 1999;9:216–221. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases