

The RNase P associated with HeLa cell mitochondria contains an essential RNA component identical in sequence to that of the nuclear RNase P
- PMID: 11134342
- PMCID: PMC86618
- DOI: 10.1128/MCB.21.2.548-561.2001
The RNase P associated with HeLa cell mitochondria contains an essential RNA component identical in sequence to that of the nuclear RNase P
Abstract
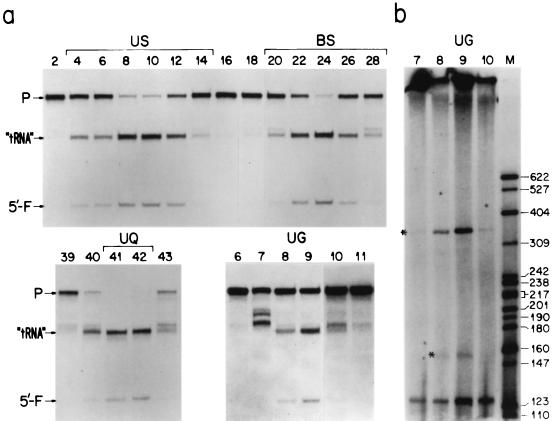
The mitochondrion-associated RNase P activity (mtRNase P) was extensively purified from HeLa cells and shown to reside in particles with a sedimentation constant ( approximately 17S) very similar to that of the nuclear enzyme (nuRNase P). Furthermore, mtRNase P, like nuRNase P, was found to process a mitochondrial tRNA(Ser(UCN)) precursor [ptRNA(Ser(UCN))] at the correct site. Treatment with micrococcal nuclease of highly purified mtRNase P confirmed earlier observations indicating the presence of an essential RNA component. Furthermore, electrophoretic analysis of 3'-end-labeled nucleic acids extracted from the peak of glycerol gradient-fractionated mtRNase P revealed the presence of a 340-nucleotide RNA component, and the full-length cDNA of this RNA was found to be identical in sequence to the H1 RNA of nuRNase P. The proportions of the cellular H1 RNA recovered in the mitochondrial fractions from HeLa cells purified by different treatments were quantified by Northern blots, corrected on the basis of the yield in the same fractions of four mitochondrial nucleic acid markers, and shown to be 2 orders of magnitude higher than the proportions of contaminating nuclear U2 and U3 RNAs. In particular, these experiments revealed that a small fraction of the cell H1 RNA (of the order of 0.1 to 0.5%), calculated to correspond to approximately 33 to approximately 175 intact molecules per cell, is intrinsically associated with mitochondria and can be removed only by treatments which destroy the integrity of the organelles. In the same experiments, the use of a probe specific for the RNA component of RNase MRP showed the presence in mitochondria of 6 to 15 molecules of this RNA per cell. The available evidence indicates that the levels of mtRNase P detected in HeLa cells should be fully adequate to satisfy the mitochondrial tRNA synthesis requirements of these cells.
Figures





Comment in
-
Difference between mitochondrial RNase P and nuclear RNase P.Mol Cell Biol. 2001 Dec;21(23):8236-7. doi: 10.1128/MCB.21.23.8236-8237.2001. Mol Cell Biol. 2001. PMID: 11710332 Free PMC article. No abstract available.
References
-
- Adhya S, Ghosh T, Das A, Bera S K, Mahaptra S. Role of an RNA-binding protein in import of tRNA into Leishmania mitochondria. J Biol Chem. 1997;272:21396–21402. - PubMed
-
- Altman S, Smith J D. Tyrosine tRNA precursor molecule polynucleotide sequence. Nat New Biol. 1971;233:35–39. - PubMed
-
- Altman S, Baer M, Guerrier-Takada C, Vioque A. Enzymatic cleavage of RNA by RNA. Trends Biochem Sci. 1986;11:515–518.
-
- Anderson S, Bankier A T, Barrell B G, de Bruijn M H L, Coulson A R, Drouin J, Eperon E I, Nierlich D P, Roe B A, Sanger F, Schreier P H, Smith A J H, Staden R, Young I G. Sequence and organization of the human mitochondrial genome. Nature. 1981;290:457–465. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources