

Tailoring in vitro evolution for protein affinity or stability
- PMID: 11134506
- PMCID: PMC14547
- DOI: 10.1073/pnas.98.1.75
Tailoring in vitro evolution for protein affinity or stability
Abstract
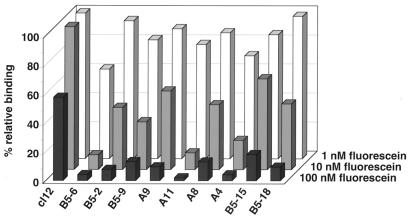
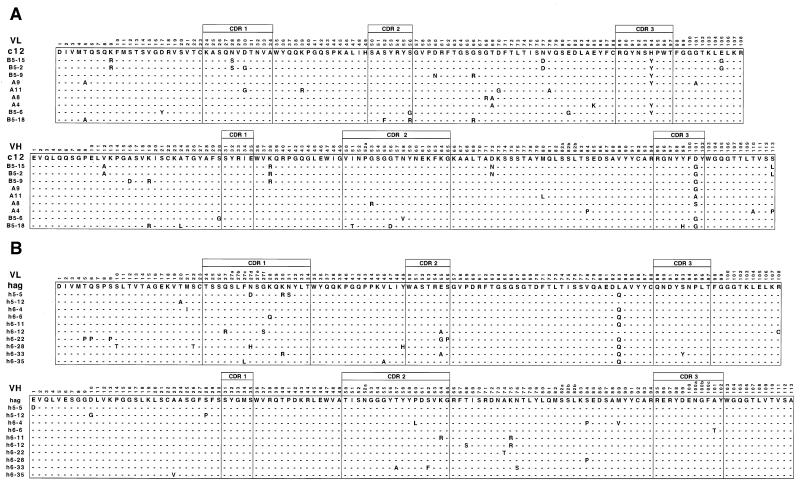
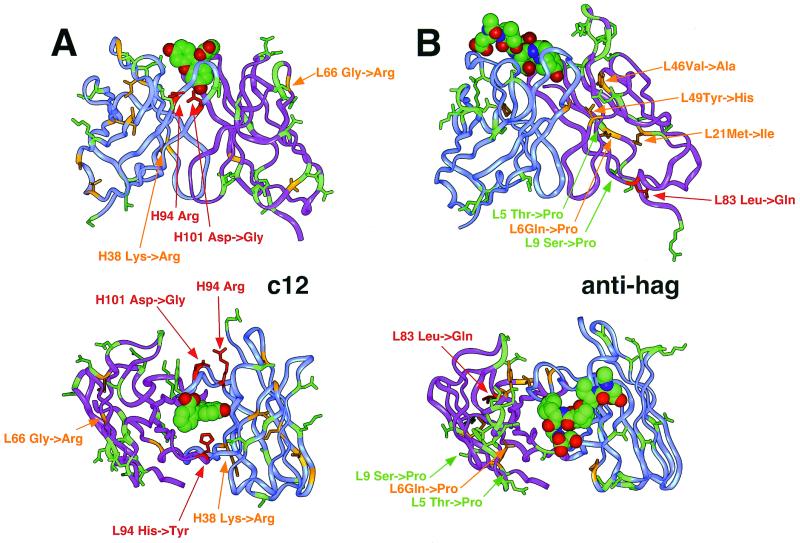
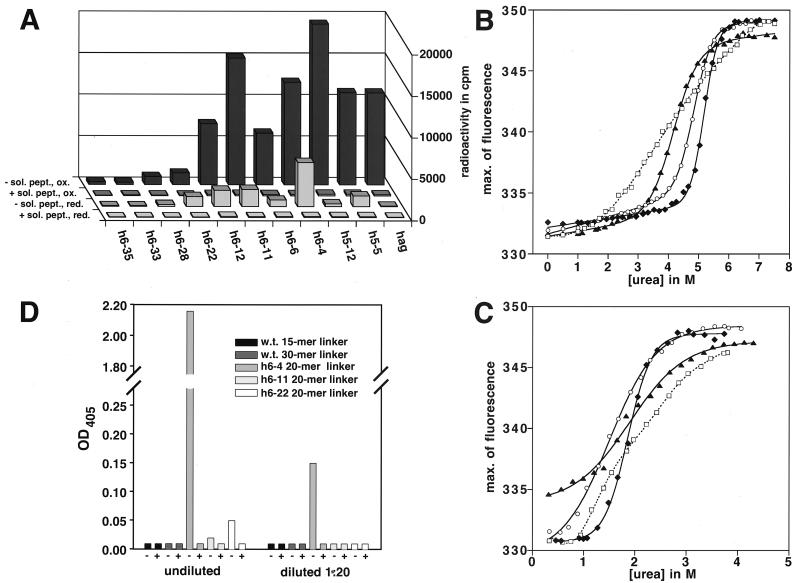
We describe a rapid and general technology working entirely in vitro to evolve either the affinity or the stability of ligand-binding proteins, depending on the chosen selection pressure. Tailored in vitro selection strategies based on ribosome display were combined with in vitro diversification by DNA shuffling to evolve either the off-rate or thermodynamic stability of single-chain Fv antibody fragments (scFvs). To demonstrate the potential of this method, we chose to optimize two proteins already possessing favorable properties. A scFv with an initial affinity of 1.1 nM (k(off) at 4 degrees C of 10(-4) s(-1)) was improved 30-fold by the use of off-rate selections over a period of several days. As a second example, a generic selection strategy for improved stability exploited the property of ribosome display that the conditions can be altered under which the folding of the displayed protein occurs. We used decreasing redox potentials in the selection step to select for molecules stable in the absence of disulfide bonds. They could be functionally expressed in the reducing cytoplasm, and, when allowed to form disulfides again, their stability had increased to 54 kJ/mol from an initial value of 24 kJ/mol. Sequencing revealed that the evolved mutant proteins had used different strategies of residue changes to adapt to the selection pressure. Therefore, by a combination of randomization and appropriate selection strategies, an in vitro evolution of protein properties in a predictable direction is possible.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
