

An amino acid code for protein folding
- PMID: 11136249
- PMCID: PMC14552
- DOI: 10.1073/pnas.98.1.105
An amino acid code for protein folding
Abstract
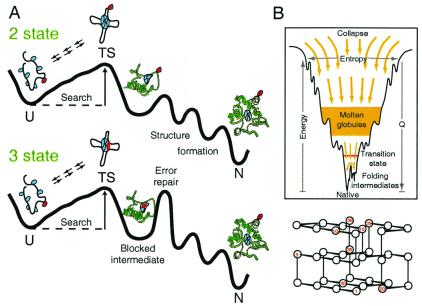
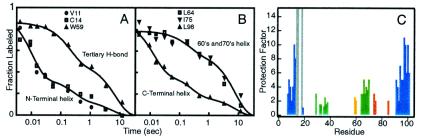
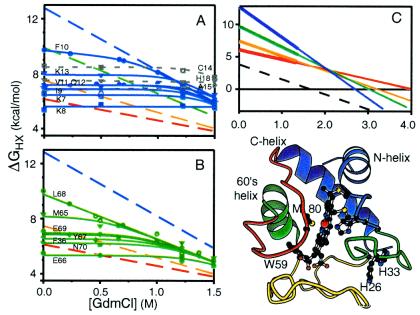
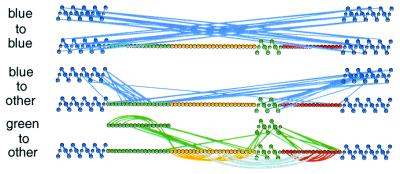
Direct structural information obtained for many proteins supports the following conclusions. The amino acid sequences of proteins can stabilize not only the final native state but also a small set of discrete partially folded native-like intermediates. Intermediates are formed in steps that use as units the cooperative secondary structural elements of the native protein. Earlier intermediates guide the addition of subsequent units in a process of sequential stabilization mediated by native-like tertiary interactions. The resulting stepwise self-assembly process automatically constructs a folding pathway, whether linear or branched. These conclusions are drawn mainly from hydrogen exchange-based methods, which can depict the structure of infinitesimally populated folding intermediates at equilibrium and kinetic intermediates with subsecond lifetimes. Other kinetic studies show that the polypeptide chain enters the folding pathway after an initial free-energy-uphill conformational search. The search culminates by finding a native-like topology that can support forward (native-like) folding in a free-energy-downhill manner. This condition automatically defines an initial transition state, the search for which sets the maximum possible (two-state) folding rate. It also extends the sequential stabilization strategy, which depends on a native-like context, to the first step in the folding process. Thus the native structure naturally generates its own folding pathway. The same amino acid code that translates into the final equilibrium native structure-by virtue of propensities, patterning, secondary structural cueing, and tertiary context-also produces its kinetic accessibility.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
