

DNA binding sites for the Mlc and NagC proteins: regulation of nagE, encoding the N-acetylglucosamine-specific transporter in Escherichia coli
- PMID: 11139621
- PMCID: PMC29661
- DOI: 10.1093/nar/29.2.506
DNA binding sites for the Mlc and NagC proteins: regulation of nagE, encoding the N-acetylglucosamine-specific transporter in Escherichia coli
Abstract
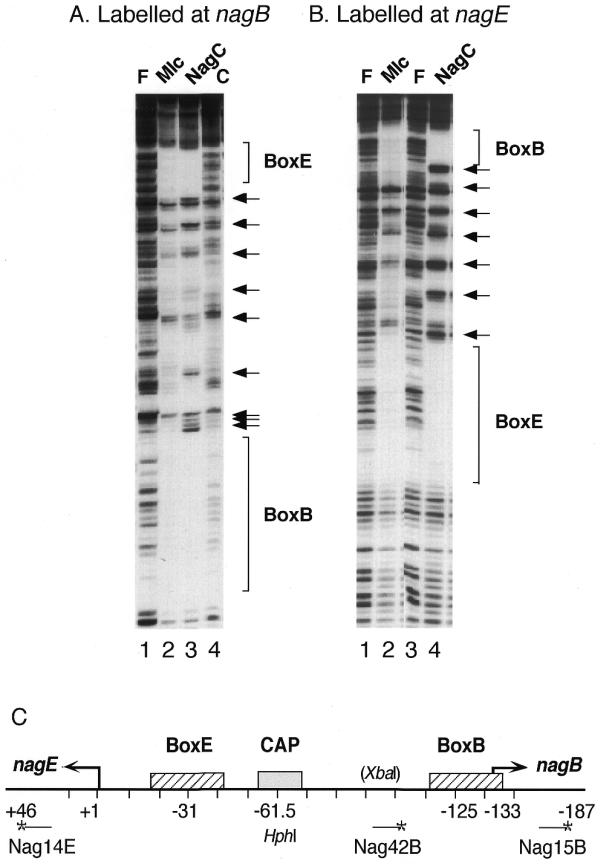
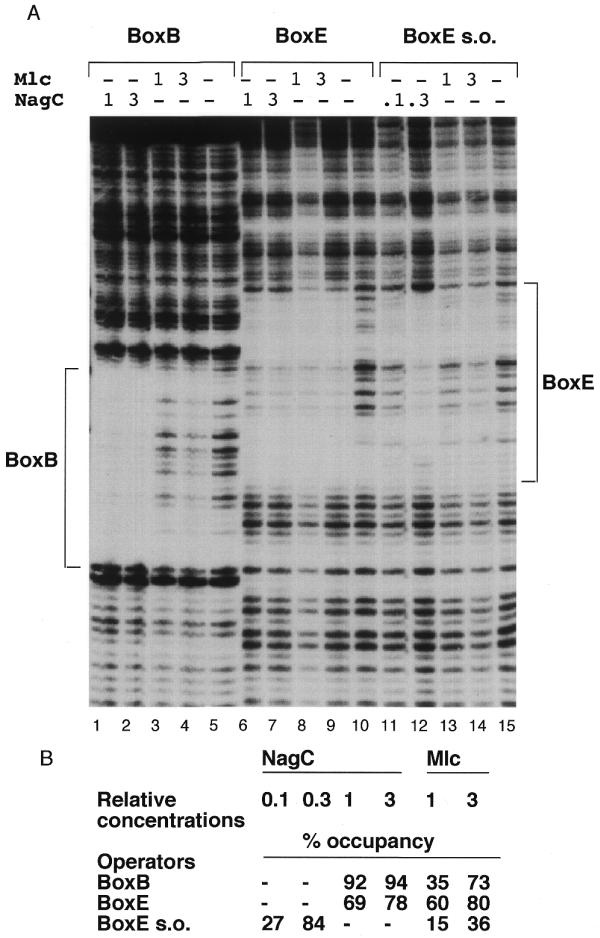
The NagC and Mlc proteins are homologous transcriptional regulators that control the expression of several phosphotransferase system (PTS) genes in Escherichia coli. NagC represses nagE, encoding the N:-acetylglucosamine-specific transporter, while Mlc represses three PTS operons, ptsG, manXYZ and ptsHIcrr, involved in the uptake of glucose. NagC and Mlc can bind to each others operator, at least in vitro. A binding site selection procedure was used to try to distinguish NagC and Mlc sites. The major difference was that all selected NagC binding sites had a G or a C at positions +11/-11 from the centre of symmetry. This is also the case for most native NagC sites, but not the nagE operator, which thus looks like a potential Mlc target. The nagE operator does exhibit a higher affinity for Mlc than NagC, but no regulation of nagE by physiological concentrations of Mlc was detected in vivo. Regulation of wild-type nagE by NagC is achieved because of the chelation effect due to a second high affinity NagC operator covering the nagB promoter. Replacing the A/T at +11/-11 with C/G allows repression by NagC in the absence of the nagB operator.
Figures


Similar articles
-
Regulation of PTS gene expression by the homologous transcriptional regulators, Mlc and NagC, in Escherichia coli (or how two similar repressors can behave differently).J Mol Microbiol Biotechnol. 2001 Jul;3(3):371-80. J Mol Microbiol Biotechnol. 2001. PMID: 11361067
-
Nag repressor-operator interactions: protein-DNA contacts cover more than two turns of the DNA helix.J Mol Biol. 1995 Jun 23;249(5):890-902. doi: 10.1006/jmbi.1995.0346. J Mol Biol. 1995. PMID: 7791215
-
Control of the expression of the manXYZ operon in Escherichia coli: Mlc is a negative regulator of the mannose PTS.Mol Microbiol. 1998 Jan;27(2):369-80. doi: 10.1046/j.1365-2958.1998.00685.x. Mol Microbiol. 1998. PMID: 9484892
-
Regulation of gene expression in the PTS in Escherichia coli: the role and interactions of Mlc.Curr Opin Microbiol. 2002 Apr;5(2):187-93. doi: 10.1016/s1369-5274(02)00296-5. Curr Opin Microbiol. 2002. PMID: 11934616 Review.
-
Transcription regulators controlled by interaction with enzyme IIB components of the phosphoenolpyruvate: sugar phosphotransferase system.Biochim Biophys Acta. 2013 Jul;1834(7):1415-24. doi: 10.1016/j.bbapap.2013.01.004. Epub 2013 Jan 11. Biochim Biophys Acta. 2013. PMID: 23318733 Review.
Cited by
-
N-acetyl-d-glucosamine induces the expression of multidrug exporter genes, mdtEF, via catabolite activation in Escherichia coli.J Bacteriol. 2006 Aug;188(16):5851-8. doi: 10.1128/JB.00301-06. J Bacteriol. 2006. PMID: 16885453 Free PMC article.
-
Switching control of expression of ptsG from the Mlc regulon to the NagC regulon.J Bacteriol. 2008 Jul;190(13):4677-86. doi: 10.1128/JB.00315-08. Epub 2008 May 9. J Bacteriol. 2008. PMID: 18469102 Free PMC article.
-
Reshaping Phosphatase Substrate Preference for Controlled Biosynthesis Using a "Design-Build-Test-Learn" Framework.Adv Sci (Weinh). 2024 Jun;11(22):e2309852. doi: 10.1002/advs.202309852. Epub 2024 Mar 19. Adv Sci (Weinh). 2024. PMID: 38504470 Free PMC article.
-
Integrated laboratory evolution and rational engineering of GalP/Glk-dependent Escherichia coli for higher yield and productivity of L-tryptophan biosynthesis.Metab Eng Commun. 2021 Feb 13;12:e00167. doi: 10.1016/j.mec.2021.e00167. eCollection 2021 Jun. Metab Eng Commun. 2021. PMID: 33665119 Free PMC article.
-
Mlc regulation of Salmonella pathogenicity island I gene expression via hilE repression.Nucleic Acids Res. 2007;35(6):1822-32. doi: 10.1093/nar/gkm060. Epub 2007 Feb 28. Nucleic Acids Res. 2007. PMID: 17329372 Free PMC article.
References
-
- Plumbridge J. and Kolb,A. (1993) DNA loop formation between Nag repressor molecules bound to its two operator sites is necessary for repression of the nag regulon of Escherichia coli in vivo. Mol. Microbiol., 10, 973–981. - PubMed
-
- Plumbridge J.A. (1991) Repression and induction of the nag regulon of Escherichia coli K12: the roles of nagC and nagA in maintenance of the uninduced state. Mol. Microbiol., 5, 2053–2062. - PubMed
-
- Hosono K., Kakuda,H. and Ichihara,S. (1995) Decreasing accumulation of acetate in rich medium by Escherichia coli on introduction of genes on a multicopy plasmid. Biosci. Biotechnol. Biochem ., 59, 256–261. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
