

The molybdenum cofactor biosynthetic protein Cnx1 complements molybdate-repairable mutants, transfers molybdenum to the metal binding pterin, and is associated with the cytoskeleton
- PMID: 11148290
- PMCID: PMC102230
- DOI: 10.1105/tpc.12.12.2455
The molybdenum cofactor biosynthetic protein Cnx1 complements molybdate-repairable mutants, transfers molybdenum to the metal binding pterin, and is associated with the cytoskeleton
Abstract
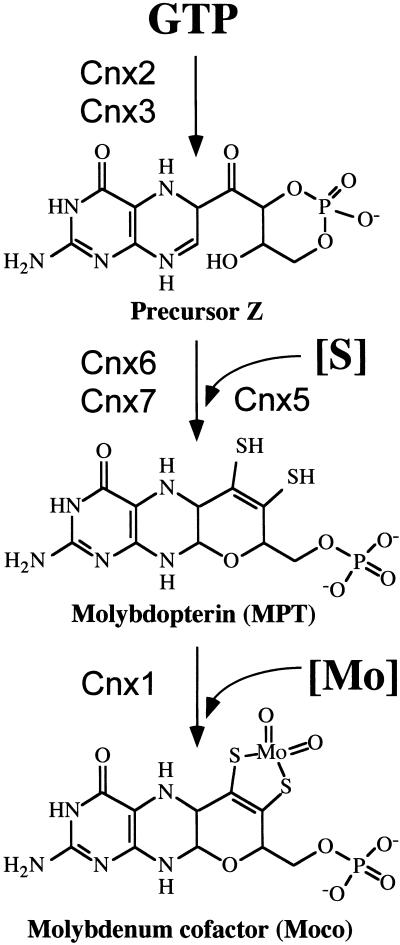
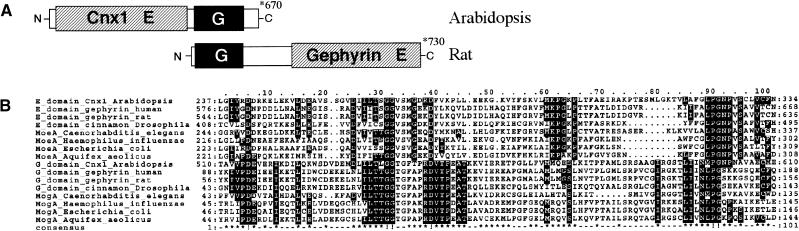
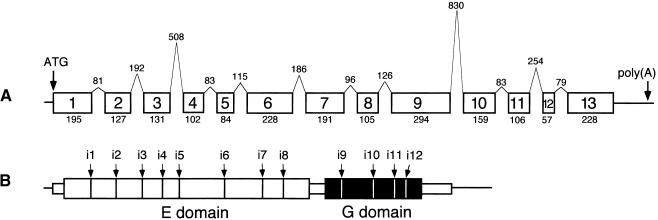
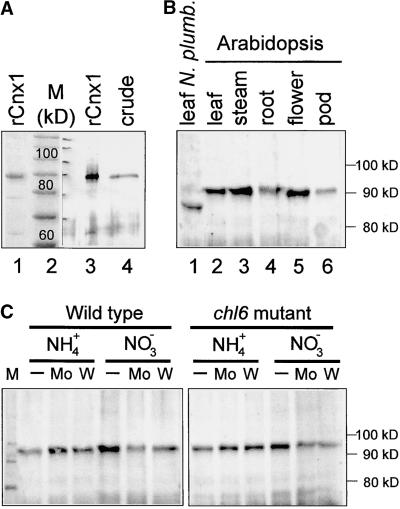
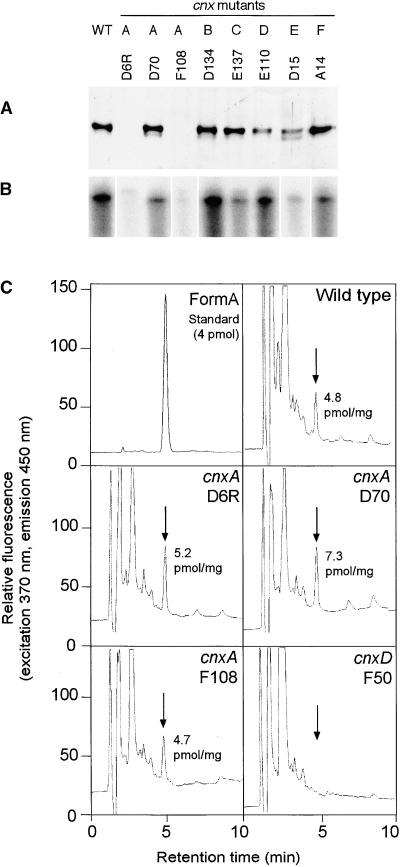
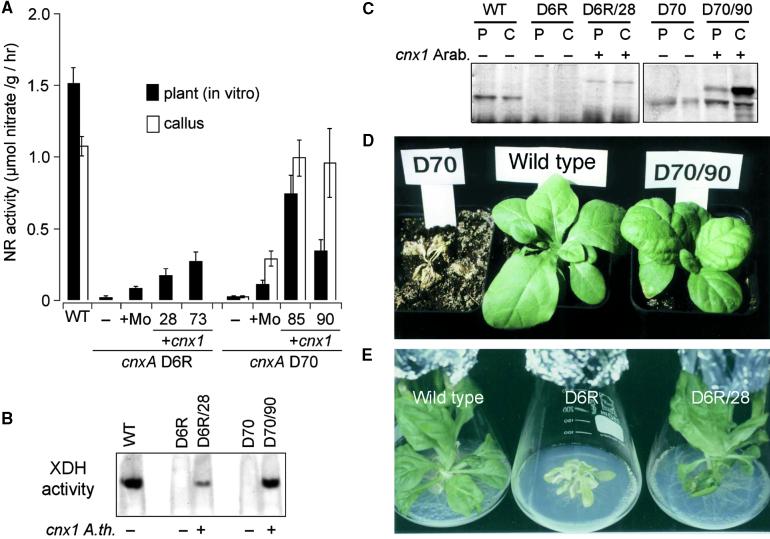
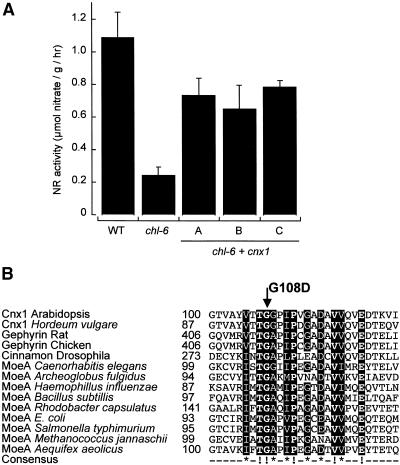
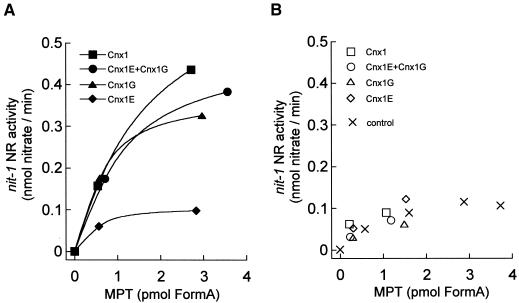
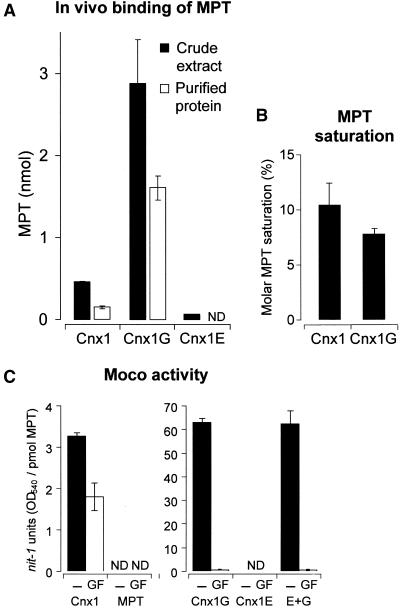
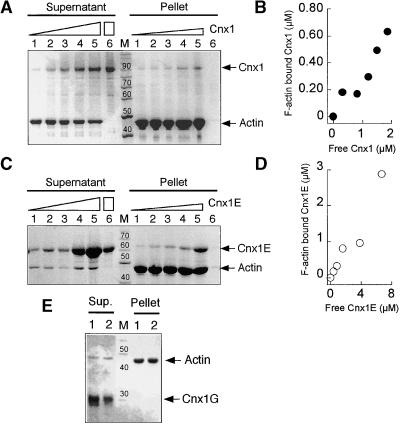
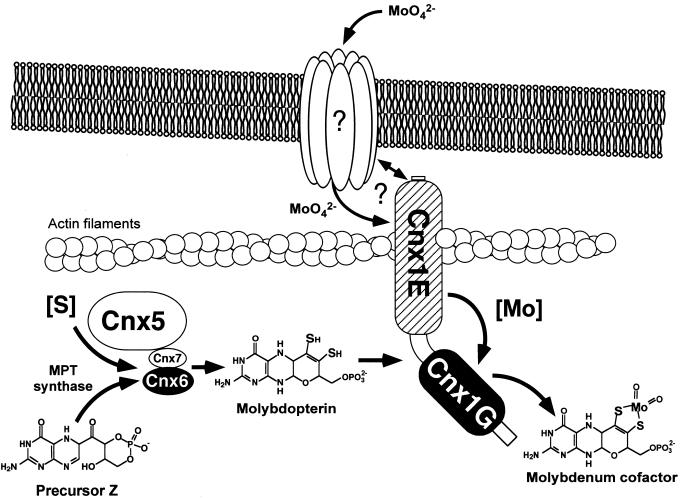
Molybdenum (Mo) plays an essential role in the active site of all eukaryotic Mo-containing enzymes. In plants, Mo enzymes are important for nitrate assimilation, phytohormone synthesis, and purine catabolism. Mo is bound to a unique metal binding pterin (molybdopterin [MPT]), thereby forming the active Mo cofactor (Moco), which is highly conserved in eukaryotes, eubacteria, and archaebacteria. Here, we describe the function of the two-domain protein Cnx1 from Arabidopsis in the final step of Moco biosynthesis. Cnx1 is constitutively expressed in all organs and in plants grown on different nitrogen sources. Mo-repairable cnxA mutants from Nicotiana plumbaginifolia accumulate MPT and show altered Cnx1 expression. Transformation of cnxA mutants and the corresponding Arabidopsis chl-6 mutant with cnx1 cDNA resulted in functional reconstitution of their Moco deficiency. We also identified a point mutation in the Cnx1 E domain of Arabidopsis chl-6 that causes the molybdate-repairable phenotype. Recombinant Cnx1 protein is capable of synthesizing Moco. The G domain binds and activates MPT, whereas the E domain is essential for activating Mo. In addition, Cnx1 binds to the cytoskeleton in the same way that its mammalian homolog gephyrin does in neuronal cells, which suggests a hypothetical model for anchoring the Moco-synthetic machinery by Cnx1 in plant cells.
Figures
References
-
- Bechtold, N., Ellis, J., and Pelletier, G. (1993). In planta Agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C.R. Acad. Sci. Paris Sci. Vie 316 1194–1199.
-
- Braaksma, F.J., and Feenstra, W.J. (1982). Isolation and characterization of nitrate reductase–deficient mutants of Arabidopsis thaliana. Theor. Appl. Genet. 64 83–90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
