

MYB-related transcription factor NtMYB2 induced by wounding and elicitors is a regulator of the tobacco retrotransposon Tto1 and defense-related genes
- PMID: 11148294
- PMCID: PMC102234
- DOI: 10.1105/tpc.12.12.2511
MYB-related transcription factor NtMYB2 induced by wounding and elicitors is a regulator of the tobacco retrotransposon Tto1 and defense-related genes
Abstract
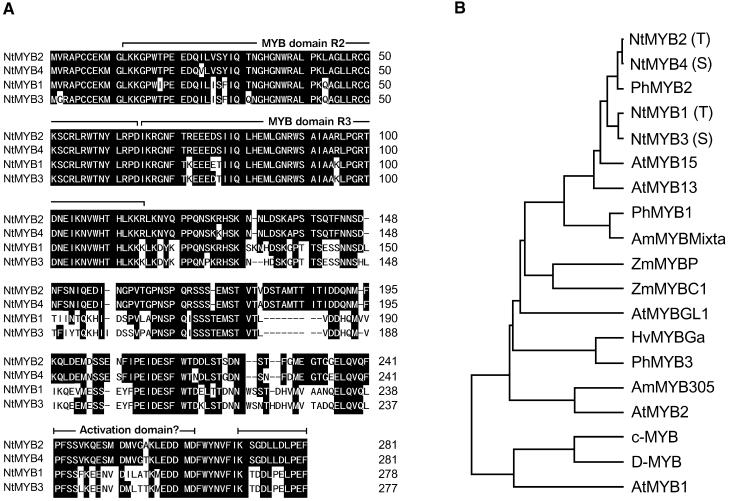
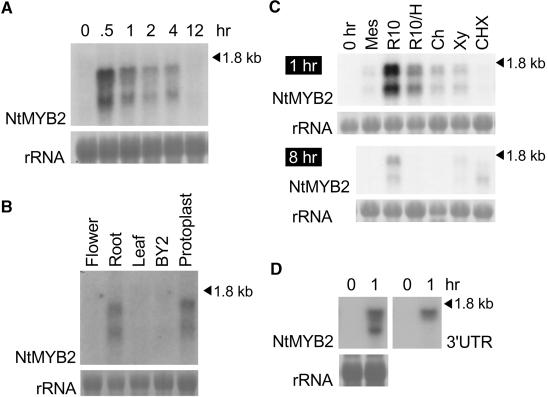
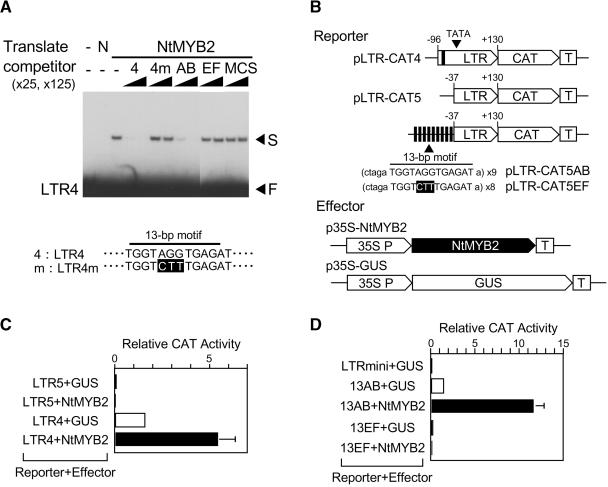
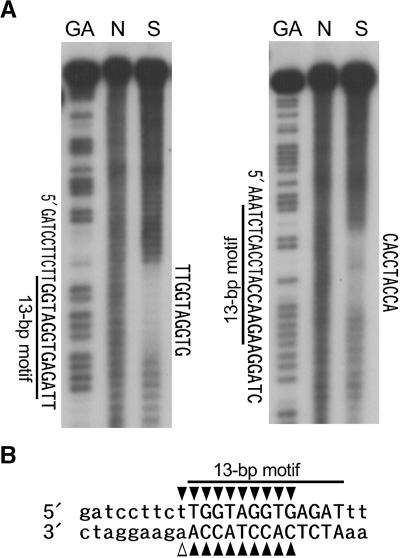
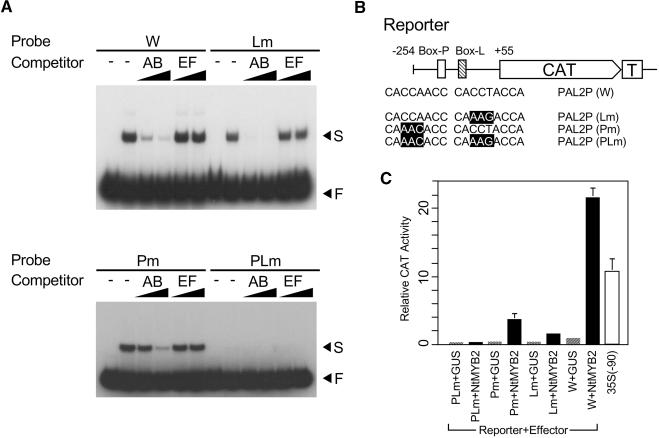
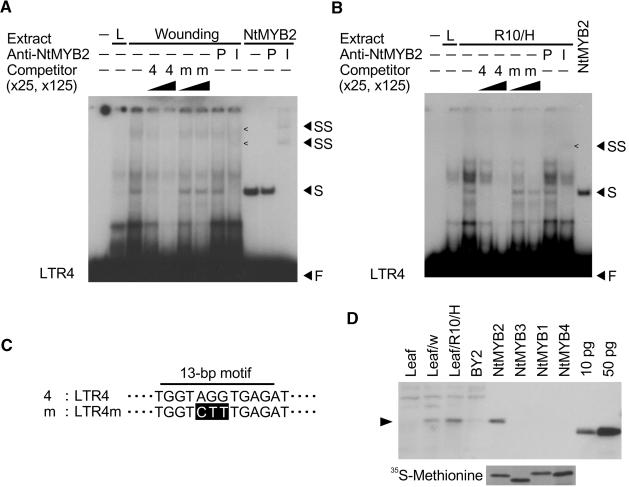
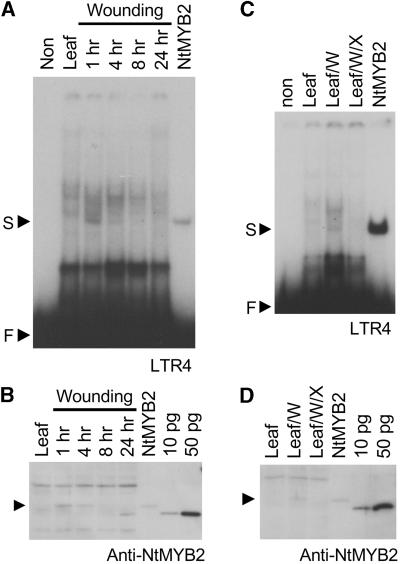
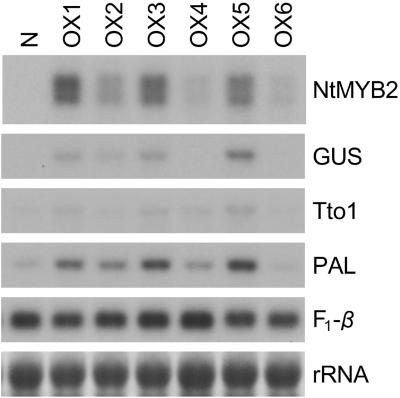
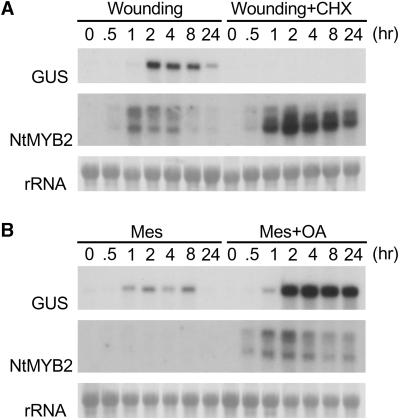
Transposition of the tobacco retrotransposon Tto1 is regulated mainly by transcription from the long terminal repeat (LTR). Functional analysis of the LTR showed that the 13-bp motif is a cis-regulatory element involved in activation by tissue culture, wounding, and treatment with elicitors. The 13-bp motif contains a conserved motif (L box) that has been implicated in the expression of phenylpropanoid synthetic genes in response to defense-related stresses. To gain further insight into the regulatory mechanism of the retrotransposon and defense-related genes, cDNAs encoding four different proteins binding to the 13-bp motif have been isolated and characterized. One protein is identical to the previously reported NtMYB1, the RNA for which is induced by virus infection; the others are also MYB-related factors. One of these factors, NtMYB2, was analyzed in detail. NtMYB2 mRNA was induced by wounding and by treatment with elicitors. NtMYB2 activated expression from the promoter with the 13-bp motif and from the promoter of the phenylalanine ammonia lyase gene (Pv-PAL2) in tobacco protoplasts. Overexpression of NtMYB2 cDNA in transgenic tobacco plants induced expression of Tto1 and a PAL gene. Together, these results indicate that NtMYB2 is involved in the stress response of the retrotransposon and defense-related genes.
Figures
Similar articles
-
Transcriptional activation mediated by binding of a plant GATA-type zinc finger protein AGP1 to the AG-motif (AGATCCAA) of the wound-inducible Myb gene NtMyb2.Plant J. 2003 Nov;36(4):550-64. doi: 10.1046/j.1365-313x.2003.01899.x. Plant J. 2003. PMID: 14617085
-
A 13-bp cis-regulatory element in the LTR promoter of the tobacco retrotransposon Tto1 is involved in responsiveness to tissue culture, wounding, methyl jasmonate and fungal elicitors.Plant J. 1999 May;18(4):383-93. doi: 10.1046/j.1365-313x.1999.00460.x. Plant J. 1999. PMID: 10406122
-
Characterization of a gene encoding a DNA-binding protein that interacts in vitro with vascular specific cis elements of the phenylalanine ammonia-lyase promoter.Plant Mol Biol. 1997 Oct;35(3):281-91. doi: 10.1023/a:1005853404242. Plant Mol Biol. 1997. PMID: 9349252
-
Functional analysis of the tobacco Tnt1 retrotransposon.Genetica. 1994;93(1-3):181-9. doi: 10.1007/BF01435250. Genetica. 1994. PMID: 7813914 Review.
-
The expression of the tobacco Tnt1 retrotransposon is linked to plant defense responses.Genetica. 1997;100(1-3):241-52. Genetica. 1997. PMID: 9440277 Review.
Cited by
-
Isolation and functional characterization of a floral tissue-specific R2R3 MYB regulator from tobacco.Planta. 2010 Apr;231(5):1061-76. doi: 10.1007/s00425-010-1108-y. Epub 2010 Feb 16. Planta. 2010. PMID: 20157728
-
Invasion of the Arabidopsis genome by the tobacco retrotransposon Tnt1 is controlled by reversible transcriptional gene silencing.Plant Physiol. 2008 Jul;147(3):1264-78. doi: 10.1104/pp.108.117846. Epub 2008 May 8. Plant Physiol. 2008. PMID: 18467467 Free PMC article.
-
Transgenerational phenotypic and epigenetic changes in response to heat stress in Arabidopsis thaliana.Plant Signal Behav. 2014;9(2):e27971. doi: 10.4161/psb.27971. Epub 2014 Feb 10. Plant Signal Behav. 2014. PMID: 24513700 Free PMC article.
-
A Genome-Wide View of the Transcriptome Dynamics of Fresh-Cut Potato Tubers.Genes (Basel). 2023 Jan 10;14(1):181. doi: 10.3390/genes14010181. Genes (Basel). 2023. PMID: 36672922 Free PMC article.
-
Oxygen supply is a prerequisite for response to aluminum in cultured cells of tobacco (Nicotiana tabacum).Plant Cell Physiol. 2025 Aug 12;66(7):1044-1060. doi: 10.1093/pcp/pcaf055. Plant Cell Physiol. 2025. PMID: 40421753 Free PMC article.
References
-
- Akama, K., Shiraishi, H., Ohta, S., Nakamura, K., Okada, K., and Shimura, Y. (1992). Efficient transformation of Arabidopsis thaliana: Comparison of the efficiencies with various organs, plant ecotypes and Agrobacterium strains. Plant Cell Rep. 12 7–11. - PubMed
-
- Avila, J., Niedo, C., Cañas, L., Benito, J., and Paz-Ares, J. (1993). Petunia hybrida genes related to the maize regulatory C1 gene and to animal myb proto-oncogenes. Plant J. 3 553–562. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
