

Replication origins in Xenopus egg extract Are 5-15 kilobases apart and are activated in clusters that fire at different times
- PMID: 11149917
- PMCID: PMC2193667
- DOI: 10.1083/jcb.152.1.15
Replication origins in Xenopus egg extract Are 5-15 kilobases apart and are activated in clusters that fire at different times
Abstract
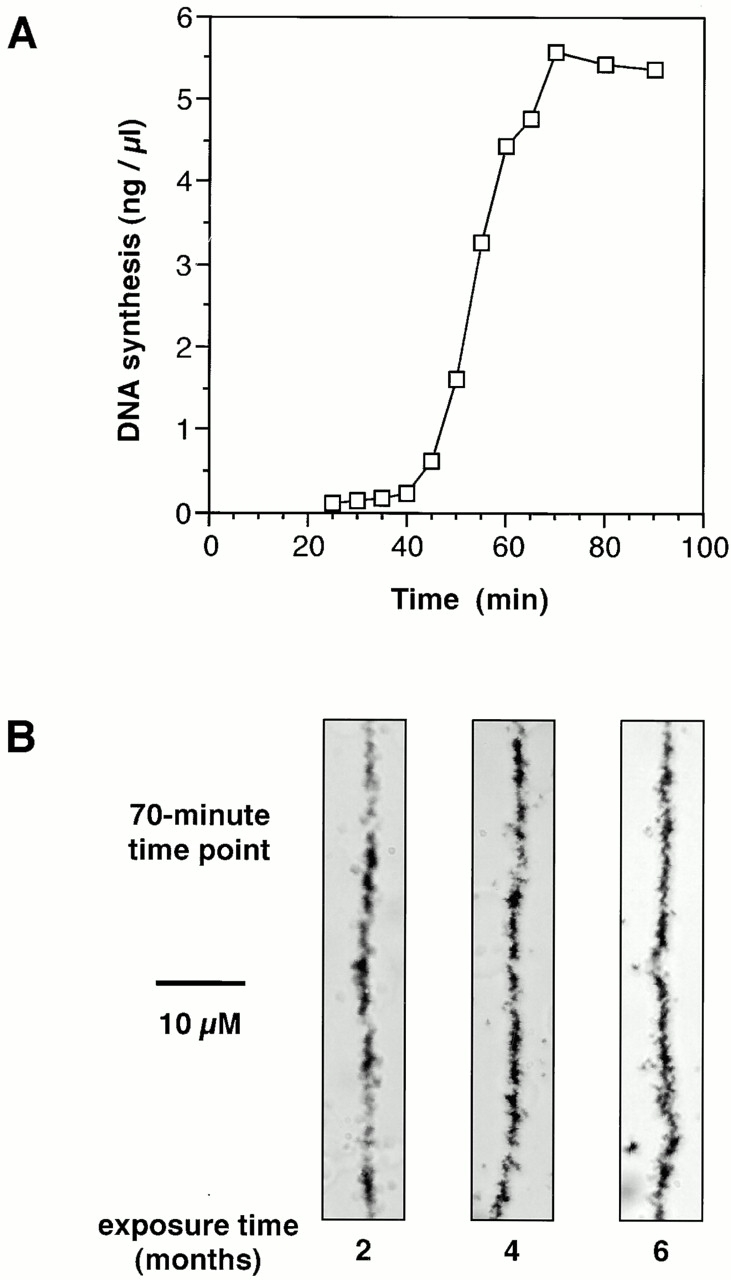
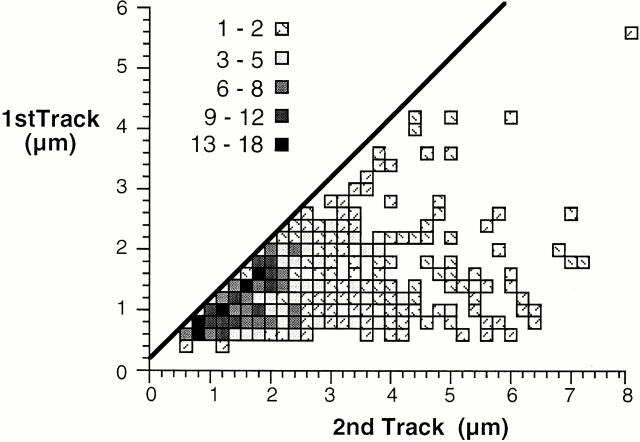
When Xenopus eggs and egg extracts replicate DNA, replication origins are positioned randomly with respect to DNA sequence. However, a completely random distribution of origins would generate some unacceptably large interorigin distances. We have investigated the distribution of replication origins in Xenopus sperm nuclei replicating in Xenopus egg extract. Replicating DNA was labeled with [(3)H]thymidine or bromodeoxyuridine and the geometry of labeled sites on spread DNA was examined. Most origins were spaced 5-15 kb apart. This regular distribution provides an explanation for how complete chromosome replication can be ensured although origins are positioned randomly with respect to DNA sequence. Origins were grouped into small clusters (typically containing 5-10 replicons) that fired at approximately the same time, with different clusters being activated at different times in S phase. This suggests that a temporal program of origin firing similar to that seen in somatic cells also exists in the Xenopus embryo. When the quantity of origin recognition complexes (ORCs) on the chromatin was restricted, the average interorigin distance increased, and the number of origins in each cluster decreased. This suggests that the binding of ORCs to chromatin determines the regular spacing of origins in this system.
Figures









References
-
- Blow J.J., Laskey R.A. Initiation of DNA replication in nuclei and purified DNA by a cell-free extract of Xenopus eggs. Cell. 1986;47:577–587. - PubMed
-
- Blow J.J., Laskey R.A. A role for the nuclear envelope in controlling DNA replication within the cell cycle. Nature. 1988;332:546–548. - PubMed
-
- Blow J.J., Sleeman A.M. Replication of purified DNA in Xenopus egg extract is dependent on nuclear assembly. J. Cell Sci. 1990;95:383–391. - PubMed
