

Calreticulin Is a receptor for nuclear export
- PMID: 11149926
- PMCID: PMC2193655
- DOI: 10.1083/jcb.152.1.127
Calreticulin Is a receptor for nuclear export
Abstract
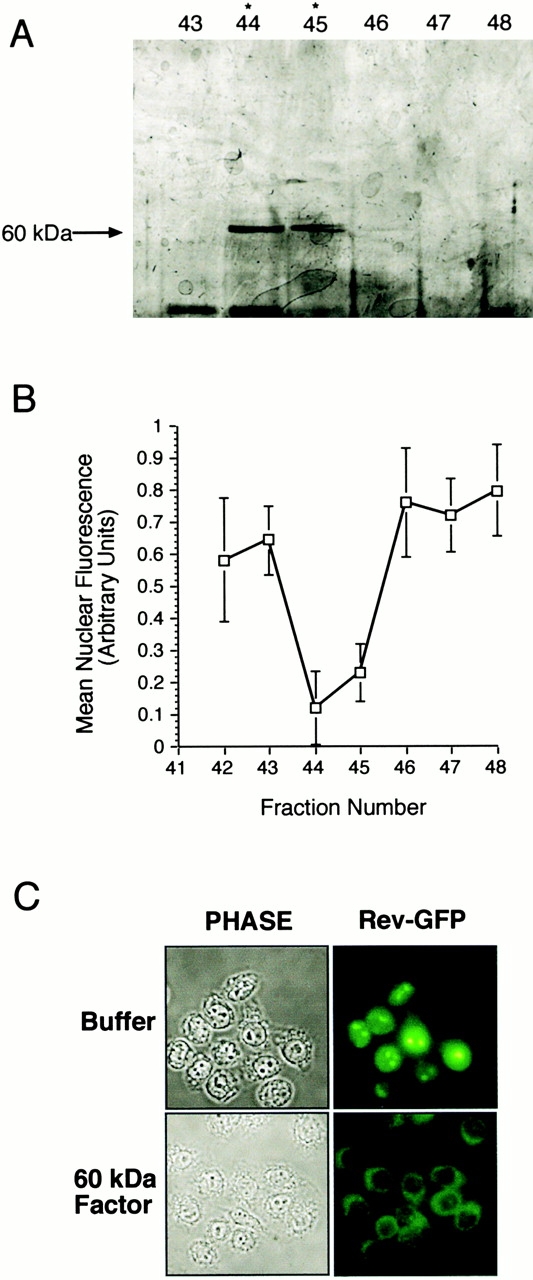
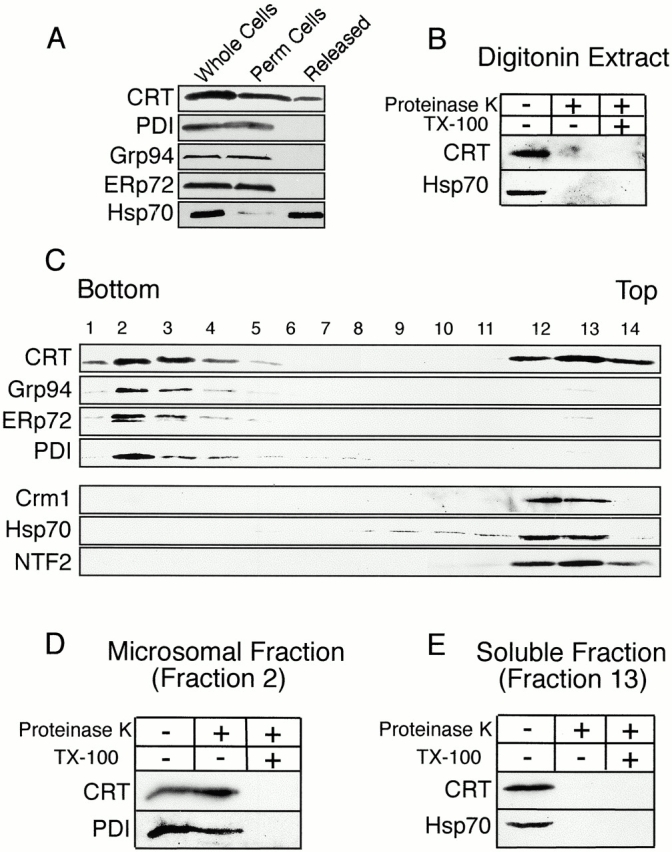
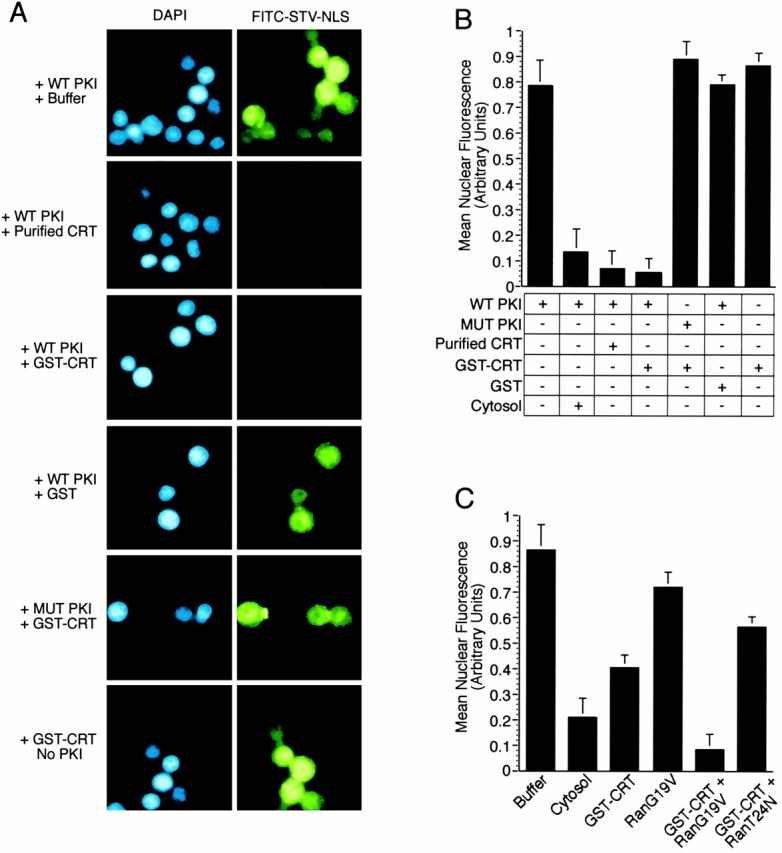
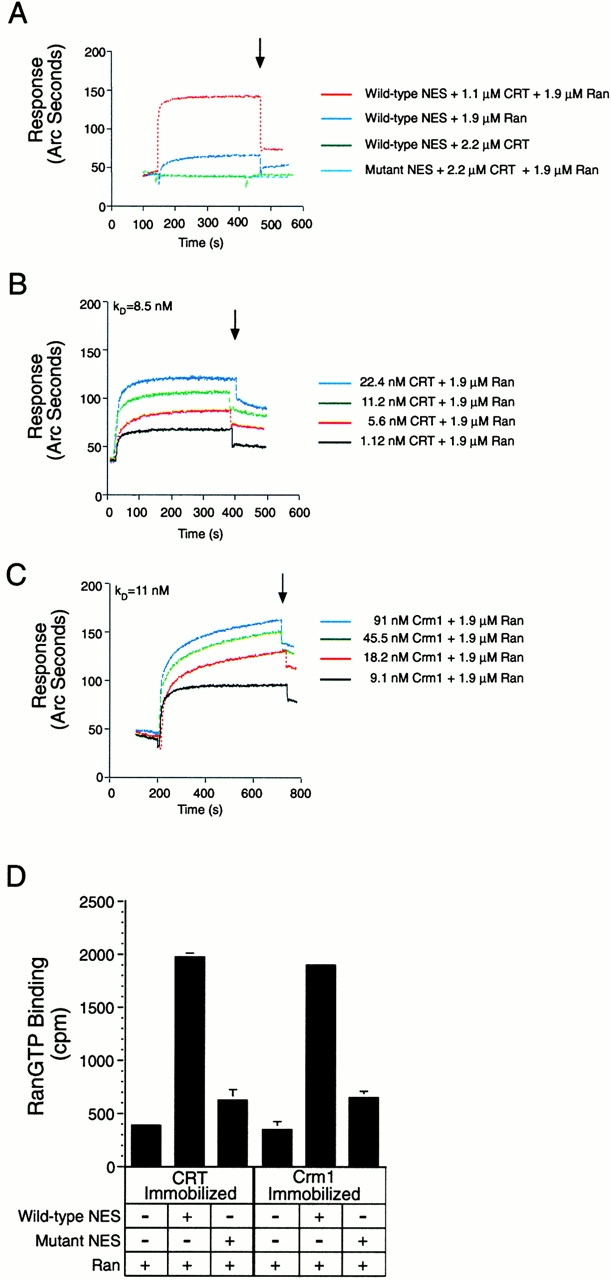
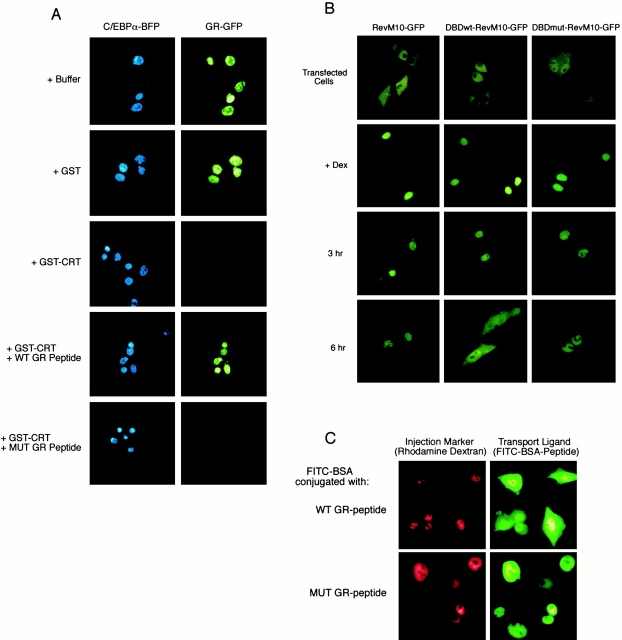
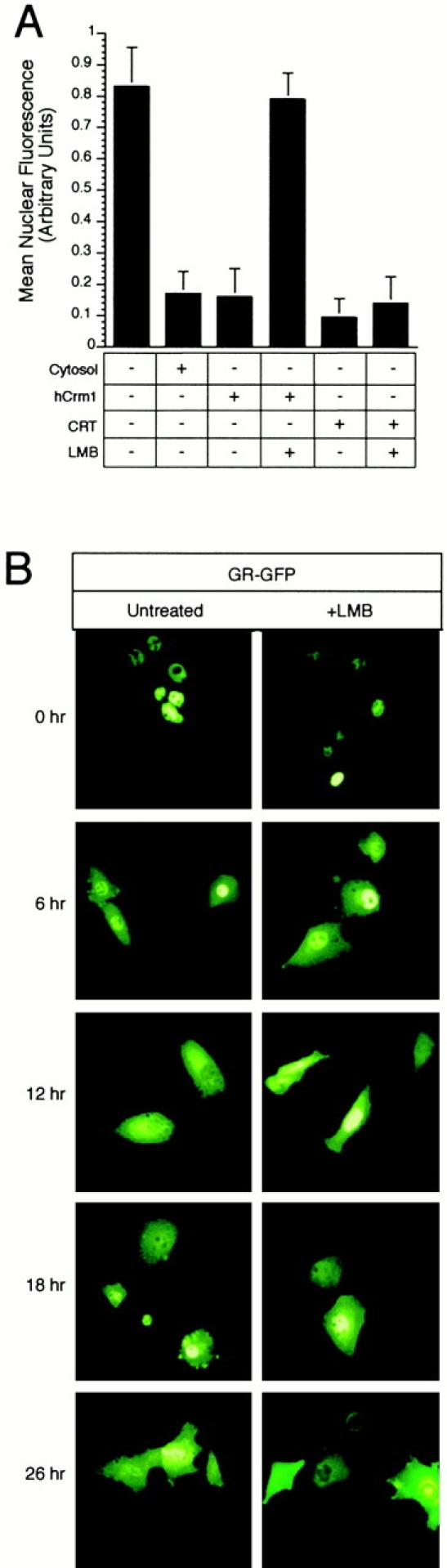
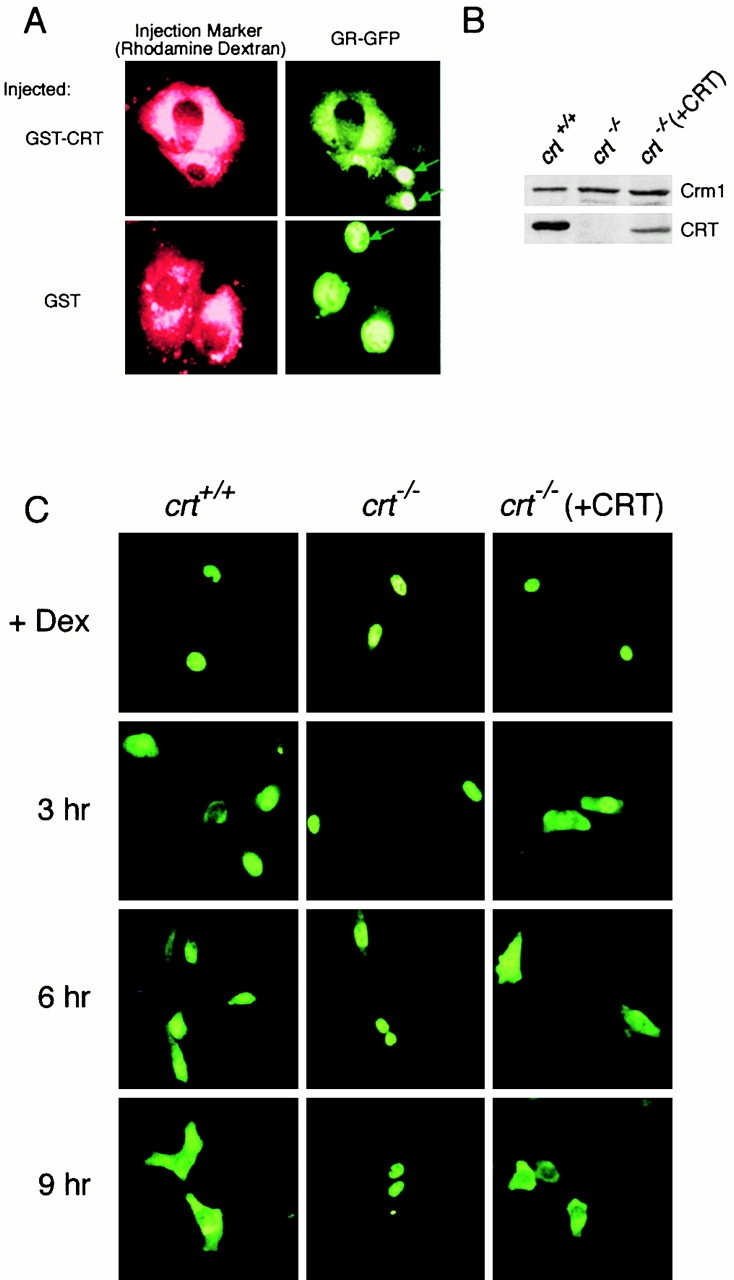
In previous work, we used a permeabilized cell assay that reconstitutes nuclear export of protein kinase inhibitor (PKI) to show that cytosol contains an export activity that is distinct from Crm1 (Holaska, J.M., and B.M. Paschal. 1995. Proc. Natl. Acad. Sci. USA. 95: 14739-14744). Here, we describe the purification and characterization of the activity as calreticulin (CRT), a protein previously ascribed to functions in the lumen of the ER. We show that cells contain both ER and cytosolic pools of CRT. The mechanism of CRT-dependent export of PKI requires a functional nuclear export signal (NES) in PKI and involves formation of an export complex that contains RanGTP. Previous studies linking CRT to downregulation of steroid hormone receptor function led us to examine its potential role in nuclear export of the glucocorticoid receptor (GR). We found that CRT mediates nuclear export of GR in permeabilized cell, microinjection, and transfection assays. GR export is insensitive to the Crm1 inhibitor leptomycin B in vivo, and it does not rely on a leucine-rich NES. Rather, GR export is facilitated by its DNA-binding domain, which is shown to function as an NES when transplanted to a green fluorescent protein reporter. CRT defines a new export pathway that may regulate the transcriptional activity of steroid hormone receptors.
Figures
References
-
- Arts G.J., Fornerod M., Mattaj I.W. Identification of a nuclear export receptor for tRNA Curr. Biol. 8 1998. 305 314a - PubMed
-
- Askjaer P., Jensen T.H., Nilsson J., Englmeier L., Kjems J. The specificity of the CRM1-Rev nuclear export signal interaction is mediated by RanGTP. J. Biol. Chem. 1998;273:33414–33422. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
