

Essential role of the small GTPase Rac in disease resistance of rice
- PMID: 11149940
- PMCID: PMC14661
- DOI: 10.1073/pnas.98.2.759
Essential role of the small GTPase Rac in disease resistance of rice
Abstract
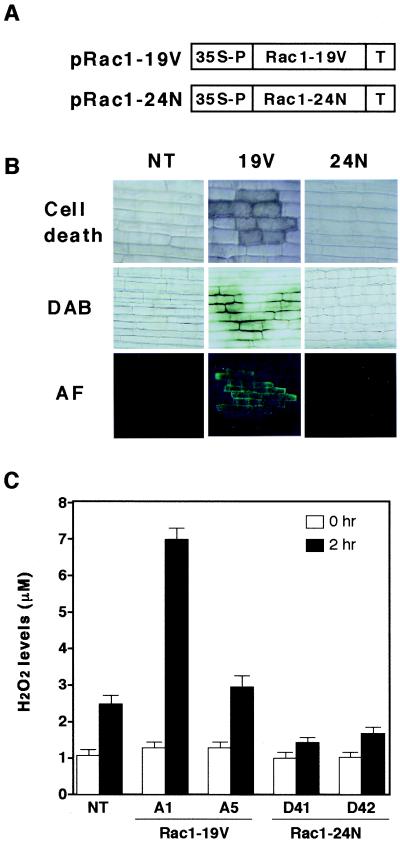
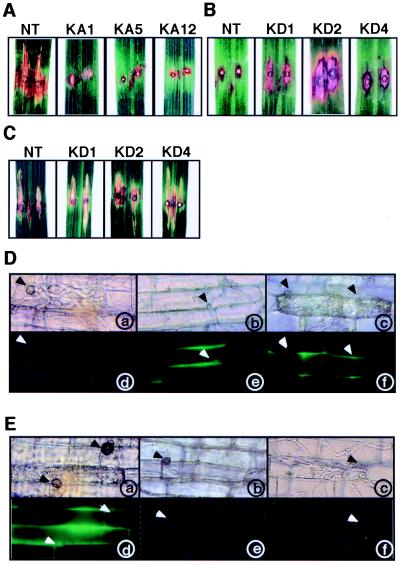
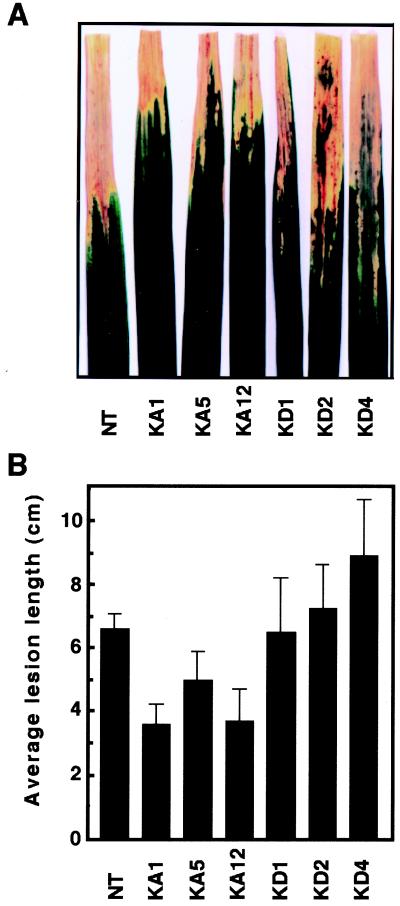
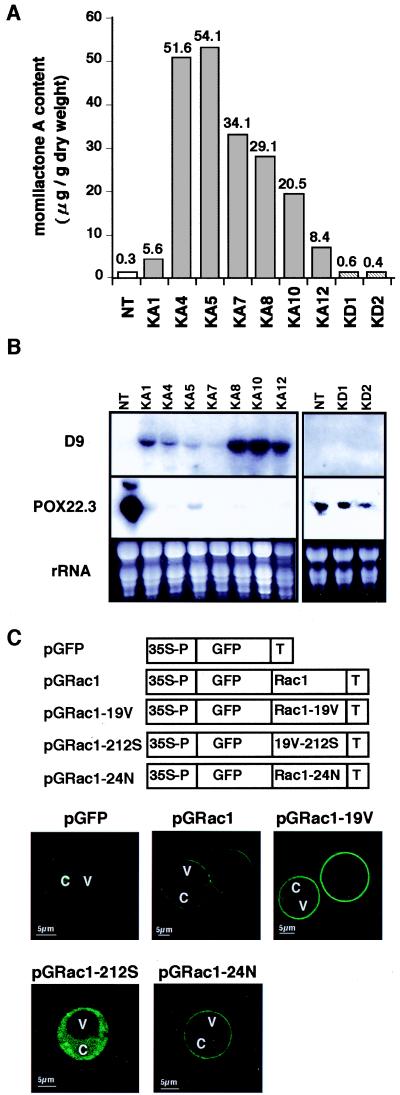
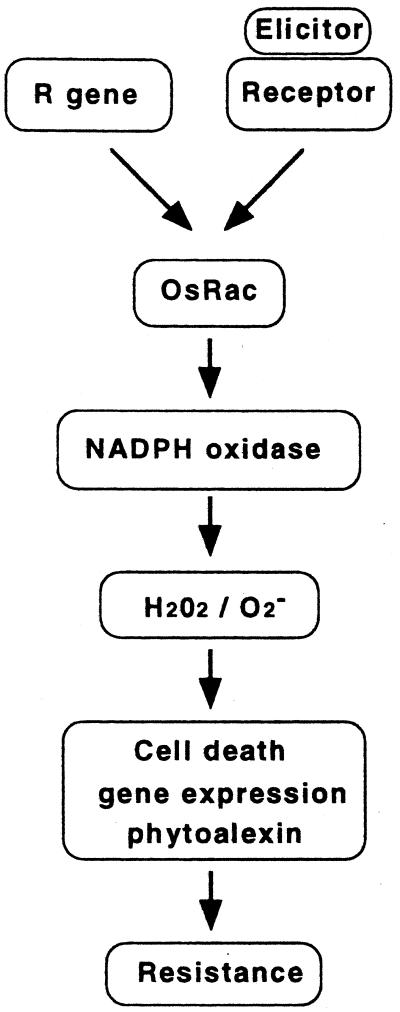
Production of reactive oxygen intermediates (ROI) and a form of programmed cell death called hypersensitive response (HR) are often associated with disease resistance of plants. We have previously shown that the Rac homolog of rice, OsRac1, is a regulator of ROI production and cell death in rice. Here we show that the constitutively active OsRac1 (i) causes HR-like responses and greatly reduces disease lesions against a virulent race of the rice blast fungus; (ii) causes resistance against a virulent race of bacterial blight; and (iii) causes enhanced production of a phytoalexin and alters expression of defense-related genes. The dominant-negative OsRac1 suppresses elicitor-induced ROI production in transgenic cell cultures, and in plants suppresses the HR induced by the avirulent race of the fungus. Taken together, our findings strongly suggest that OsRac1 has a general role in disease resistance of rice.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
