

Comparative Study
doi: 10.1073/pnas.98.2.468.
Epub 2001 Jan 9.
Stepwise unfolding of titin under force-clamp atomic force microscopy
Affiliations
- PMID: 11149943
- PMCID: PMC14610
- DOI: 10.1073/pnas.98.2.468
Item in Clipboard
Comparative Study
Stepwise unfolding of titin under force-clamp atomic force microscopy
Proc Natl Acad Sci U S A.
.
Abstract
Here we demonstrate the implementation of a single-molecule force clamp adapted for use with an atomic force microscope. We show that under force-clamp conditions, an engineered titin protein elongates in steps because of the unfolding of its modules and that the waiting times to unfold are exponentially distributed. Force-clamp measurements directly measure the force dependence of the unfolding probability and readily captures the different mechanical stability of the I27 and I28 modules of human cardiac titin. Force-clamp spectroscopy promises to be a direct way to probe the mechanical stability of elastic proteins such as those found in muscle, the extracellular matrix, and cell adhesion.
Figures
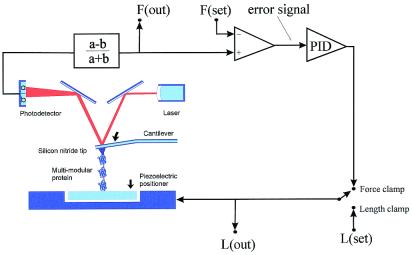
Schematic diagram of the AFM apparatus and associated control
electronics. When pressed against a layer of protein attached to a
substrate, the silicon nitride tip of the AFM cantilever may adsorb a
single protein molecule. Extension of the molecule by retraction of the
piezoelectric positioner results in deflection of the cantilever. We
can stretch a single molecule by using either a length-clamp mode or a
force-clamp mode. In the standard length-clamp mode we set the desired
position (L) and measure the resulting force (F) calculated from the
laser deflection (a − b)/(a + b). In the force-clamp mode, the
measured force is compared with a set value generating an error signal
that is fed to a proportional, integral and differential amplifier
(PID) whose output is connected directly to the piezoelectric
positioner.
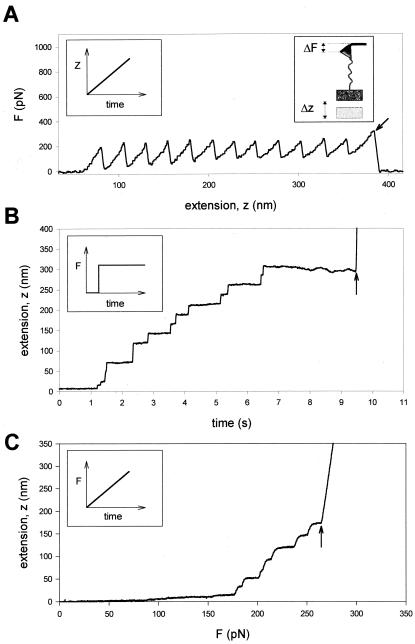
Comparison of single protein unfolding events captured with an AFM in
length-clamp mode (A) and a force-clamp mode
(B and C). (A) In standard
mode AFM, the positioner is moved linearly as a function of time
(Left Inset) and the pulling force is measured from the
degree of deflection of the cantilever (Right Inset).
When a modular protein is stretched between the cantilever tip and a
coverslip attached to a piezo-electric positioner, the resulting
force-extension curve shows a sawtooth pattern with equally spaced
force peaks, where each force peak represents the unfolding of single
protein domains. The recording corresponds to the stretching and
unfolding of a polyprotein containing 12 identical repeats of titin Ig
domain I27. The last peak corresponds to the detachment of the protein
from the cantilever and the measured force drops to zero (arrow).
(B and C) In constant force AFM, the
force applied on the protein is kept at a set value by using a
force-clamp system that continuously changes the position of the
piezoelectric device. (B) When a step increase in force
is applied to a single I27 polyprotein, the extension-time curve shows
step increases in the length of the protein, where each step is about
22 nm in size. This trace was obtained by stepping the force from −400
to 180 pN. (C) Under force-clamp conditions the force
applied to a single I2712 polyprotein is increased linearly
with time. The figure shows the resulting length versus force trace. As
shown, stepwise unfolding events occur over a narrow range of forces
(in this case ≈170–250 pN) and a much shorter time (≈2 s)
than those observed under constant force. In these experiments the
steps are slightly rounded because of the limited frequency response of
the feedback system. In both cases (B and
C), the last step increase in length marks the
detachment of the protein from the cantilever, where the position of
the piezoelectric actuator increases rapidly to saturation (arrows).
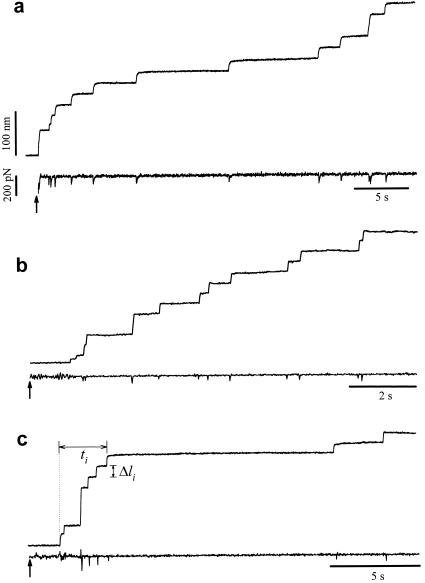
Different examples of protein unfolding events captured with
constant-force AFM. In these examples, the set point was stepped from
−500 pN (cantilever pressing on the protein sample) to 200 pN
(a), 182 pN (b), and 52 pN
(c) at the times marked by the arrows. These recordings
contain between nine (c) and 12 (a and
b) unfolding events that occur in a time interval of 9
(b) and 32 s (a and
c). Notice that some steps are two or three times the
unitary size of 22 nm. The force trace shows brief spikes that mark the
sudden unfolding of domains. These spikes have a duration of ≈20 ms
and correspond to the time response of the force clamp.
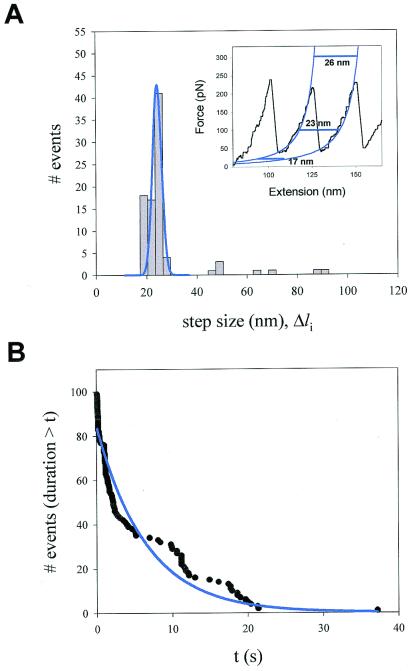
Analysis of the step sizes and dwell times of unfolding events recorded
at constant force. (A) Frequency histogram of step sizes
for the I2712 polyprotein (n = 99)
shows a principal peak centered at 22.4 nm and three additional minor
peaks that average 48.1, 67.3, and 89.1 nm. The data were obtained from
16 different experiments at an average force of 147 pN.
(Inset) Fits (blue lines) of the worm-like chain (Eq.
1) to a sawtooth pattern of an I2712
polyprotein. The horizontal lines illustrate the expected length
increase caused by domain unfolding at various forces. (B)
Probability distribution function showing the probability that after
applying force, a module remains folded for a time equal or less than
t. The distribution was fit by a single exponential function
with a time constant τ = 6.7 s (solid line).
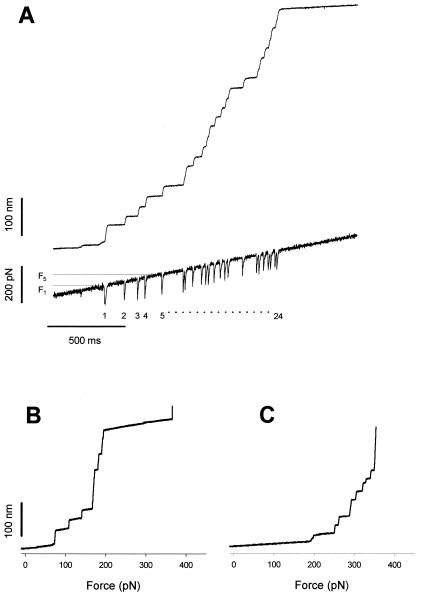
Measurements of the force dependence of the unfolding probability using
a force ramp. Force-ramp experiments apply a linearly increasing force
to a folded protein causing the full unfolding of its modules under
well-defined conditions and over a relatively short period.
(A) Stepwise unfolding of a native cardiac titin
molecule using the force-ramp method. The upper trace shows that the
molecule elongates in 24 steps over a ≈200 pN range. The lower trace
shows the time course of the force. The downward transients are caused
by the feedback lag and serve as useful markers of unfolding.
(B) Stepwise unfolding events for the I2712
polyprotein recorded under conditions similar to those shown in
A. The step increases in length are plotted as a
function of the applied force. (C) A similar experiment
done with the I288 polyprotein shows that this polyprotein
unfolds in a higher force range than the I2712
polyprotein.
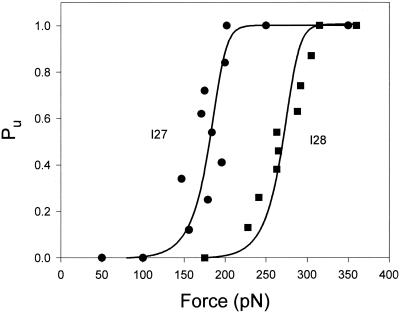
Unfolding probability (Pu) as a function of
the applied force for the I2712 polyprotein
(●) and the I288 polyprotein
(■). Pu is calculated by counting
the number of unfolding events that have occurred up to a given force.
For example, there are 24 unfolding events observed in Fig.
5A. The occurrence of event 12 marks the force at which
Pu = 0.5. ●
correspond to data obtained from the engineered I2712
polyprotein (49 steps from six experiments). ■ correspond to
data obtained from the engineered I288 polyprotein (31
steps from four experiments). Both sets of data were fitted with a
simple two-state model of unfolding that considers that the applied
force increases linearly with time (solid lines; see text).
References
-
- Fisher T E, Oberhauser A F, Carrion-Vazquez M, Marszalek P E, Fernandez J M. Trends Biochem Sci. 1999;24:379–384. - PubMed
-
- Rief M, Gautel M, Oesterhelt F, Fernandez J M, Gaub H E. Science. 1997;276:1109–1112. - PubMed
-
- Marszalek P E, Lu H, Li H B, Carrion-Vazquez M, Oberhauser A F, Schulten K, Fernandez J M. Nature (London) 1999;402:100–103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
