

Mutations in hepatitis C virus RNAs conferring cell culture adaptation
- PMID: 11152517
- PMCID: PMC114050
- DOI: 10.1128/JVI.75.3.1437-1449.2001
Mutations in hepatitis C virus RNAs conferring cell culture adaptation
Abstract
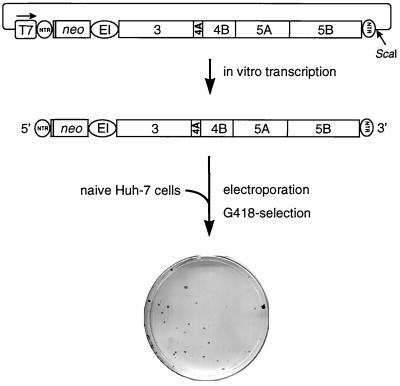
As an initial approach to studying the molecular replication mechanisms of hepatitis C virus (HCV), a major causative agent of acute and chronic liver disease, we have recently developed selectable self-replicating RNAs. These replicons lacked the region encoding the structural proteins and instead carried the gene encoding the neomycin phosphotransferase. Although the replication levels of these RNAs within selected cells were high, the number of G418-resistant colonies was reproducibly low. In a search for the reason, we performed a detailed analysis of replicating HCV RNAs and identified several adaptive mutations enhancing the efficiency of colony formation by several orders of magnitude. Adaptive mutations were found in nearly every nonstructural protein but not in the 5' or 3' nontranslated regions. The most drastic effect was found with a single-amino-acid substitution in NS5B, increasing the number of colonies approximately 500-fold. This mutation was conserved with RNAs isolated from one cell line, in contrast to other amino acid substitutions enhancing the efficiency of colony formation to a much lesser extent. Interestingly, some combinations of these nonconserved mutations with the highly adaptive one reduced the efficiency of colony formation drastically, suggesting that some adaptive mutations are not compatible.
Figures
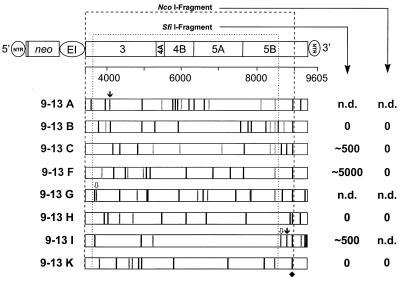
 , frameshift mutation. The single conserved amino acid substitution in NS5B is marked with a diamond. The positions of the recognition sequences for the restriction enzymes SfiI and NcoI used for subcloning into the parental construct are indicated with dotted lines. The names of the corresponding constructs are given at the left, and the CFU/μg in vitro transcript obtained with each of the subcloned NcoI or SfiI fragments after selection with G418 (500 μg/ml) is given on the right. n.d., not determined.
, frameshift mutation. The single conserved amino acid substitution in NS5B is marked with a diamond. The positions of the recognition sequences for the restriction enzymes SfiI and NcoI used for subcloning into the parental construct are indicated with dotted lines. The names of the corresponding constructs are given at the left, and the CFU/μg in vitro transcript obtained with each of the subcloned NcoI or SfiI fragments after selection with G418 (500 μg/ml) is given on the right. n.d., not determined.


References
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K, editors. Current protocols in molecular biology. New York, N.Y: John Wiley and Sons; 1987.
-
- Bartenschlager R, Lohmann V. Replication of hepatitis C virus. J Gen Virol. 2000;81:1631–1648. - PubMed
-
- Beard M R, Abell G, Honda M, Carroll A, Gartland M, Clarke B, Suzuki K, Lanford R, Sangar D V, Lemon S M. An infectious molecular clone of a Japanese genotype 1b hepatitis C virus. Hepatology. 1999;30:316–324. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
