

Genetic evidence of an essential role for cytomegalovirus small capsid protein in viral growth
- PMID: 11152518
- PMCID: PMC114051
- DOI: 10.1128/JVI.75.3.1450-1458.2001
Genetic evidence of an essential role for cytomegalovirus small capsid protein in viral growth
Abstract
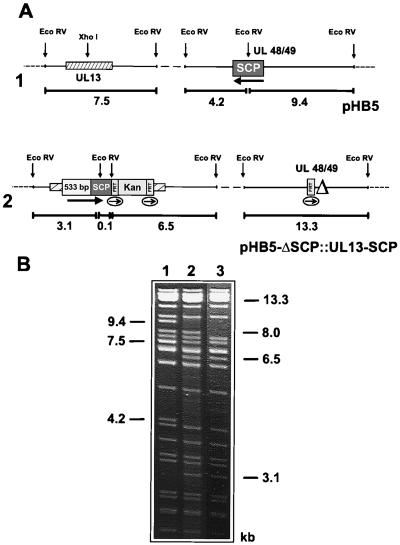
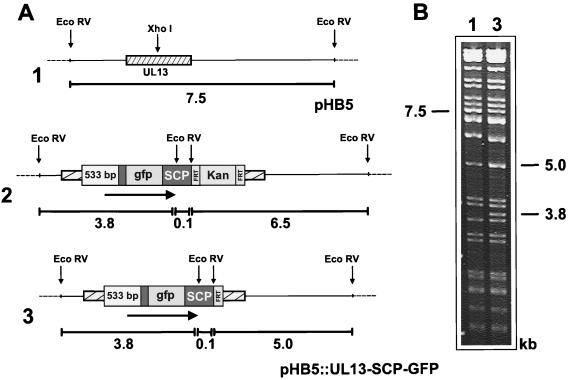
Many steps in the replication cycle of cytomegalovirus (CMV), like cell entry, capsid assembly, and egress of newly synthesized virions, have not been completely analyzed yet. In order to facilitate these studies, we decided to construct a recombinant CMV that incorporates the green fluorescent protein (GFP) into the nucleocapsid. A comparable herpes simplex virus type 1 (HSV-1) mutant has recently been generated by fusion of the GFP open reading frame (ORF) with the HSV-1 ORF encoding small capsid protein (SCP) VP26 (P. Desai and S. Person, J. Virol. 72:7563-7568, 1998). Recombinant CMV genomes expressing a fusion protein consisting of GFP and the SCP were constructed by the recently established bacterial artificial chromosome mutagenesis procedure. In transfected cells, the SCP-GFP fusion protein localized to distinct foci in the nucleus that may represent sites for capsid assembly (assemblons). However, no viable progeny was reconstituted from these mutant CMV genomes. CMV genomes with deletion of the SCP ORF also did not give rise to infectious virus. Rescue of the mutation by insertion of the SCP gene at an ectopic position in an SCP knockout genome indicates that, in contrast to the HSV-1 SCP, the CMV SCP is essential for viral growth. Expression of the SCP-GFP fusion protein together with the authentic SCP blocked the CMV infection cycle, suggesting that the SCP-GFP fusion protein exerts a dominant-negative effect on the assembly of new virions. The results of this study are discussed with regard to recently published data about the structure of the CMV virion and its differences from the HSV-1 virion.
Figures




References
-
- Borst E M, Messerle M. Development of a cytomegalovirus vector for somatic gene therapy. Bone Marrow Transplant. 2000;25(Suppl. 2):S80–S82. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
