

Review
doi: 10.1128/MCB.21.3.691-702.2001.
The Max network gone mad
Affiliations
- PMID: 11154257
- PMCID: PMC86661
- DOI: 10.1128/MCB.21.3.691-702.2001
Item in Clipboard
Review
The Max network gone mad
Mol Cell Biol.
2001 Feb.
No abstract available
Figures
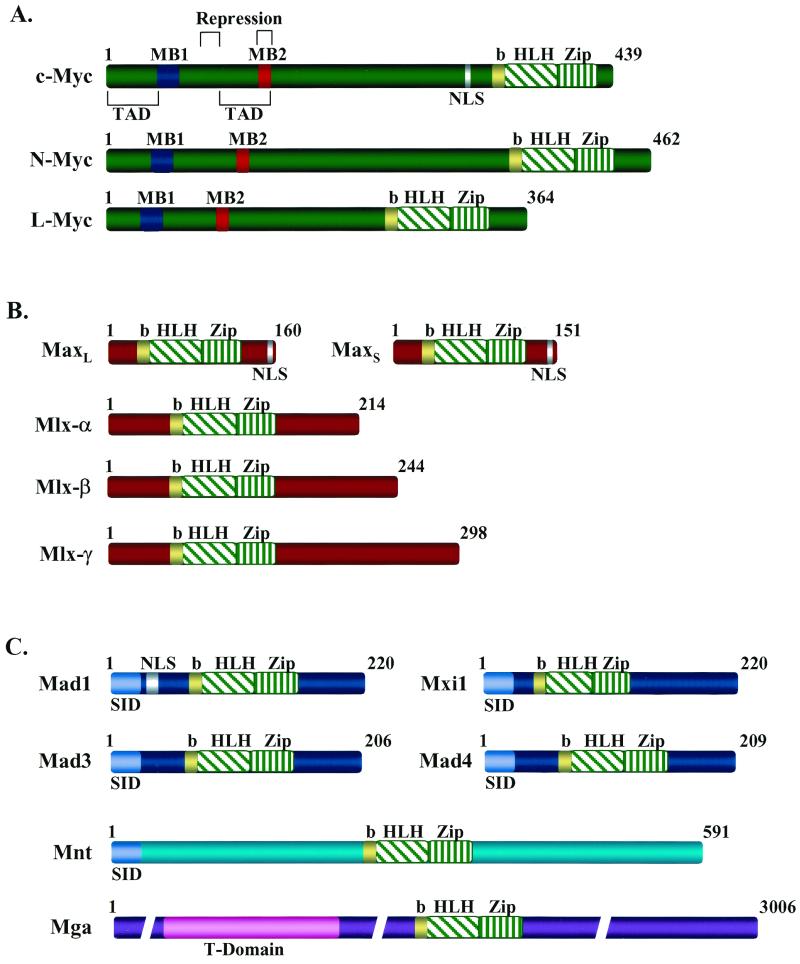
Domain structure of Myc, Max, and Mad family members and of Mnt and Mga. The structures of the bHLHZip factors of the Myc-Max-Mad network are shown. (A) The Myc family members activated in human cancer (c-Myc, N-Myc, and L-Myc). MB1 and MB2 are conserved Myc homology boxes found in all Myc family members that are required for Myc's transactivation and/or transrepression functions (113). MB2 is also required for interaction with the TRRAP transcriptional coactivator. TAD, c-Myc transactivation domain. (B) Max and its relative Mlx, which is expressed as three isoforms (18, 21, 85, 91). Max can homodimerize or form heterodimers with all members of the network through the HLHZip interactions, and the basic region confers specific DNA binding to CACGTG elements. Mlx selectively dimerizes with Mad1, Mad4, and Mnt but fails to dimerize with either Myc or Max (18, 91). (C) The Myc antagonists Mad1, Mxi1, Mad3, Mad4, and the related proteins Mnt (Rox) and Mga. Mad family members and Mnt (Rox) contain an alpha-helical amino-terminal domain (SID) that is required for interactions with the transcriptional corepressors Sin3a and Sin3b. By contrast, Mga contains a T domain near its amino terminus which is a conserved DNA- and protein-binding motif found in the Brachyury family of cell specification transcription factors (64). NLS indicates the location of identified nuclear localization signals. b, basic.
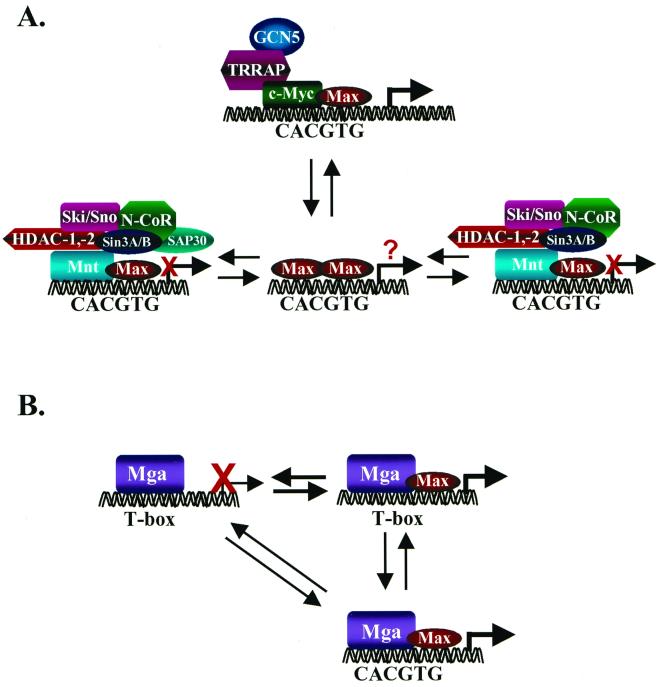
The conventional model of transcriptional regulation by the Myc-Max-Mad network (A) and transcriptional regulation by Mga (B). (A) As proposed by Eisenman and colleagues (11, 75), transcriptional activation is mediated exclusively by Myc-Max complexes which bind to CACGTG elements (and also to CACATG) in target genes. Myc's ability to activate transcription appears to require its association with the transcriptional coactivator TRRAP (89), which tethers the histone acetylase GCN5 (90). Transcriptional repression is mediated through the binding of Mad-Max or Mnt-Max complexes to identical sites and to the transcriptional corepressors Sin3a and Sin3b, through the SID. Sin3a/Sin3b interactions tether Mad/Mnt-Max complexes to a large transcriptional repressor complex that appears to contain the corepressors N-CoR, Ski, and Sno; an adapter protein, SAP30 (77, 145); and the histone deacetylases 1 and 2. An equilibrium among the various components of this network, regulated by changes in their expression and/or signaling events, dictates whether there is transcriptional activation or repression of target genes. The precise function of Max homodimers is not resolved, although they clearly exist in cells (142) and have been suggested to passively repress transcription by competition for DNA-binding elements (5). (B) Functions of Mga. Mga contains two DNA-binding protein interaction motifs, a bHLHZip domain related to that present in Myc, Max, Mad, and Mnt, and a T domain, which facilitates binding to large recognition elements that are bound by the Brachyury family of transcription factors, which play an essential role in cell specification (64). When bound to T boxes, Mga functions as a transcriptional repressor, but this is relieved by overexpression of Max. Binding of Mga to the consensus Myc-Max binding site CACGTG requires Max interactions, and at least when both are overexpressed, this complex activates gene expression (64). Thus, in theory, Mga can compete with Myc by sequestering Max, but the net result may be that Mga induces the same targets as does Myc.
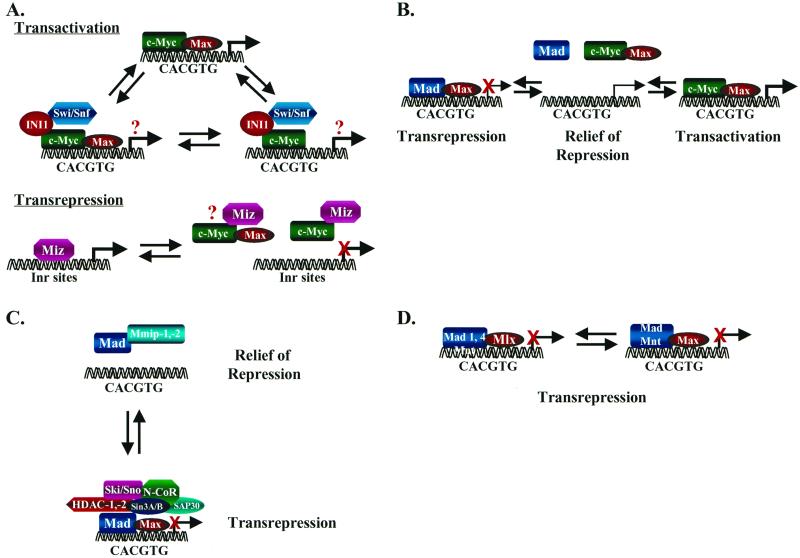
Alternative mechanisms regulating Myc transcriptional activity. (A) Myc interacts through its HLH domain, perhaps in a mutually exclusive fashion, with the transcriptional regulatory proteins Max (top), INI1/Snf5 (middle), and Miz1 (bottom) to activate (Max and INI1) or repress (Miz1) transcription. It is unknown whether ternary complexes form. (B) Target genes bearing CACGTG elements are actively repressed by Mad/Mnt-Max complexes. Myc may then relieve active repression by effectively sequestering Max into solution and/or may bind CACGTG elements in conjunction with Max to activate transcription. (C) The Mad-specific interacting proteins Mmip-1 and Mmip-2 relieve Mad-mediated active repression by sequestering Mad into solution, which may lead to transcriptional activation. Mad–Mmip-1 or Mad–Mmip-2 complexes apparently do not bind DNA (49, 143). (D) The Max-related protein Mlx selectively binds to Mad1, Mad4, and Mnt, and these complexes actively repress transcription through binding to CACGTG elements (18, 91). Thus, Mad-mediated repression of transcription can occur independently of Max.
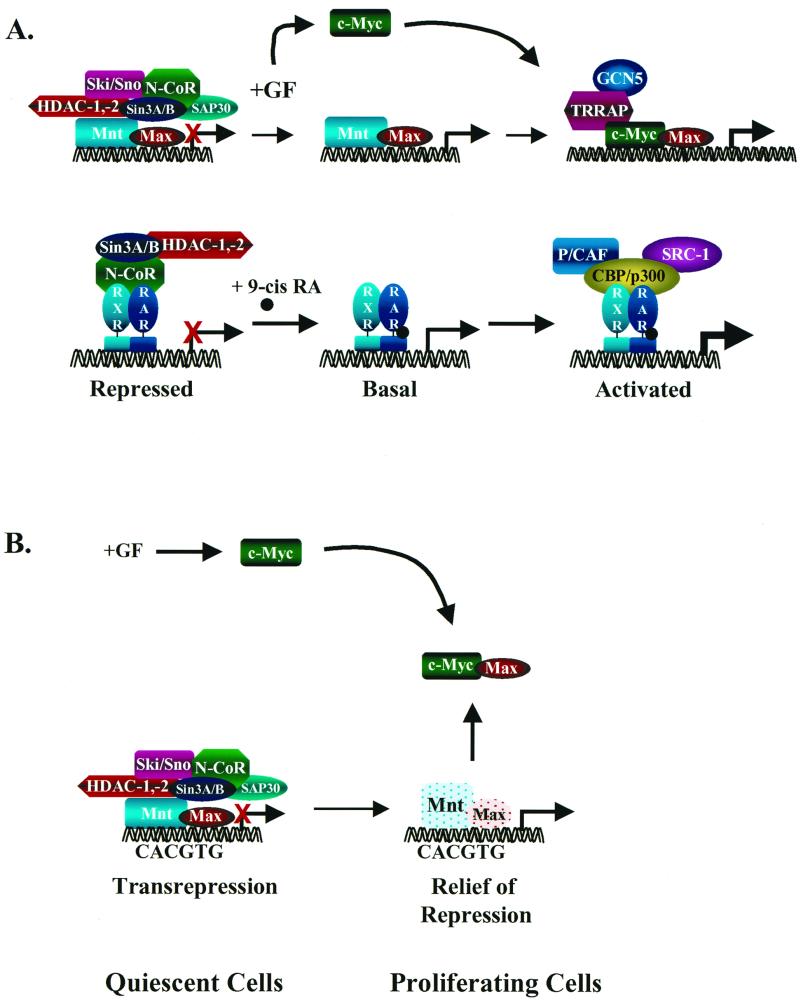
Transcriptional regulation by displacement-recruitment versus sequestration. (A) At top is shown a model of transcriptional regulation of Myc target genes by displacement and recruitment. In the repressed state, Mad/Mnt-Max DNA-binding complexes tether the transcription repressor complex. Following the addition of mitogens, c-Myc expression is up-regulated and the corepressor complex dissociates from Mad/Mnt-Max complexes, relieving active repression. c-Myc then complexes with Max, binds DNA, and recruits the transcriptional coactivator TRRAP, which tethers histone acetyltransferase GCN5. This complex then activates transcription through chromatin remodeling. This model has many parallels to the regulation of transcription by nuclear receptors (bottom panel) such as the retinoic acid receptor and retinoid X receptor (87). (B) Myc target genes are activated indirectly by the sequestration of Max away from Mad or Mnt. In this model, Myc-Max complexes do not bind DNA and the activation of target genes is due to relief of repression. Targets are then activated in proliferating cells simply by virtue of inducing c-Myc expression.
References
-
- Adams J M, Harris A W, Pinkert C A, Corcoran L M, Alexander W S, Palmiter R D, Brinster R L. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature. 1985;318:533–538. - PubMed
-
- Alitalo K, Koskinen P, Makela T P, Saksela K, Sistonen L, Winqvist R. myc oncogenes: activation and amplification. Biochim Biophys Acta. 1987;907:1–32. - PubMed
-
- Alland L, Muhle R, Hou H, Jr, Potes J, Chin L, Schreiber-Agus N, DePinho R A. Role for N-CoR and histone deacetylase in Sin3-mediated transcriptional repression. Nature. 1997;387:49–55. - PubMed
-
- Amati B, Brooks M W, Levy N, Littlewood T D, Evan G I, Land H. Oncogenic activity of the c-Myc protein requires dimerization with Max. Cell. 1993;72:233–245. - PubMed
-
- Amati B, Dalton S, Brooks M W, Littlewood T D, Evan G I, Land H. Transcriptional activation by the human c-Myc oncoprotein in yeast requires interaction with Max. Nature. 1992;359:423–426. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources