

A florigenic effect of sucrose in Fuchsia hybrida is blocked by gibberellin-induced assimilate competition
- PMID: 11154356
- PMCID: PMC61029
- DOI: 10.1104/pp.125.1.488
A florigenic effect of sucrose in Fuchsia hybrida is blocked by gibberellin-induced assimilate competition
Abstract
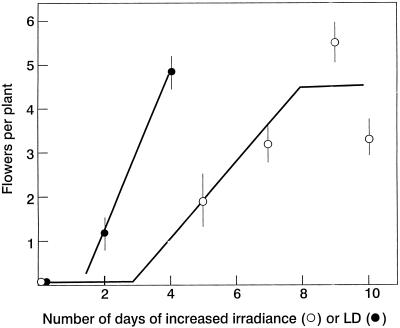
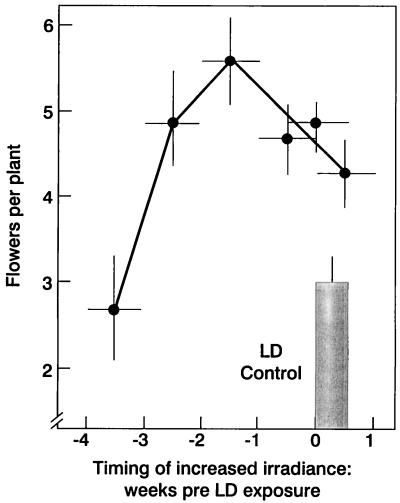
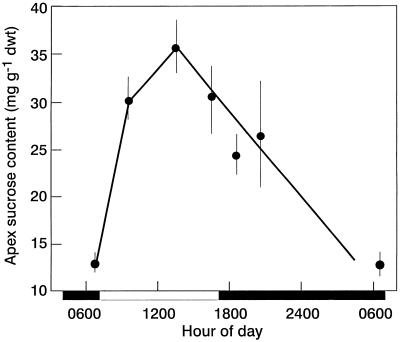
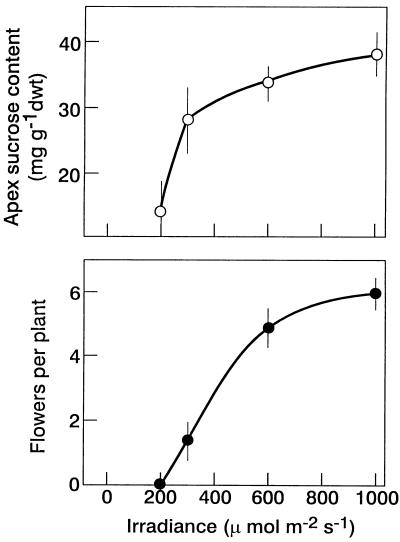
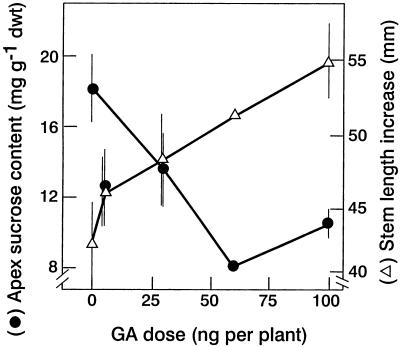
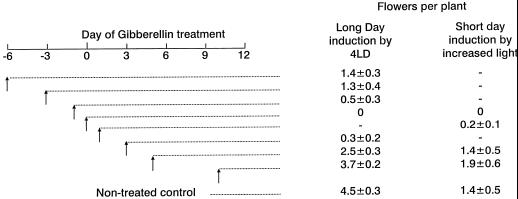
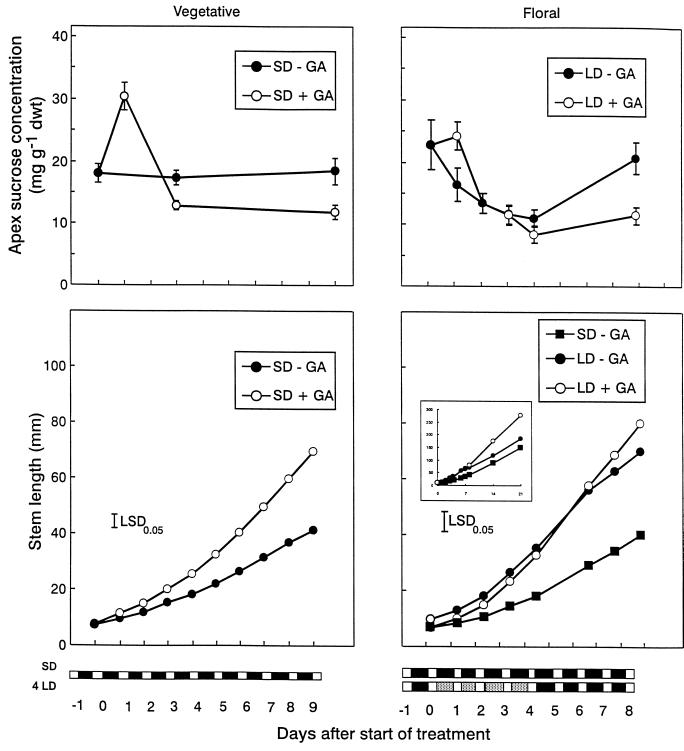
The use of gas chromatography-mass spectrometry-selected ion monitoring along with a (13)C internal standard has allowed sensitive measurements of the sucrose (Suc) content of individual shoot apices of Fuchsia hybrida. With intact plants, as the photosynthetic irradiance increased, so did shoot apex Suc content, reaching saturation at about 500 micromol m(-2) s(-1). These same plants flowered at the higher irradiances, remaining vegetative in 10-h short days at an irradiance of 230 micromol m(-2) s(-1). The strong correlation (r = 0.93) in these studies between flowering and shoot apex Suc content indicates a role for Suc as a stimulus to flowering in this species. However, Suc is not the long-day (LD) "florigen" of F. hybrida because 2 to 4 LD given as a 14-h low-irradiance photoperiod extension (10-15 micromol m(-2) s(-1)) induced flowering but without increase in shoot apex Suc content. Flowering induced by either pathway, the LD- or the Suc-mediated one, was inhibited by applying gibberellin (GA) to the shoot tip. Such inhibition of flowering by GA, at least for the LD pathway, was associated with a reduced apex Suc content, enhanced elongation of subapical stem tissue, and a reduced import into the shoot apex of leaf-sourced assimilate. Thus, our findings show how GA inhibits flowering of F. hybrida and confirm the importance of nutrient diversion in regulating flowering.
Figures
Similar articles
-
Long-day induction of flowering in Lolium temulentum involves sequential increases in specific gibberellins at the shoot apex.Plant Physiol. 2001 Oct;127(2):624-32. Plant Physiol. 2001. PMID: 11598236 Free PMC article.
-
Synthesis of gibberellin GA6 and its role in flowering of Lolium temulentum.Phytochemistry. 2003 Jan;62(1):77-82. doi: 10.1016/s0031-9422(02)00447-8. Phytochemistry. 2003. PMID: 12475622
-
Selective deactivation of gibberellins below the shoot apex is critical to flowering but not to stem elongation of Lolium.Mol Plant. 2008 Mar;1(2):295-307. doi: 10.1093/mp/ssm030. Epub 2008 Feb 8. Mol Plant. 2008. PMID: 19825541
-
Gibberellins and flowering of grasses and cereals: prizing open the lid of the "florigen" black box.Annu Rev Plant Biol. 2003;54:307-28. doi: 10.1146/annurev.arplant.54.031902.135029. Annu Rev Plant Biol. 2003. PMID: 14502993 Review.
-
Universal florigenic signals triggered by FT homologues regulate growth and flowering cycles in perennial day-neutral tomato.J Exp Bot. 2006;57(13):3405-14. doi: 10.1093/jxb/erl106. Epub 2006 Sep 27. J Exp Bot. 2006. PMID: 17005925 Review.
Cited by
-
The nature of floral signals in Arabidopsis. I. Photosynthesis and a far-red photoresponse independently regulate flowering by increasing expression of FLOWERING LOCUS T (FT).J Exp Bot. 2008;59(14):3811-20. doi: 10.1093/jxb/ern231. Epub 2008 Oct 3. J Exp Bot. 2008. PMID: 18836142 Free PMC article.
-
SlCAND1, encoding cullin-associated Nedd8-dissociated protein 1, regulates plant height, flowering time, seed germination, and root architecture in tomato.Plant Mol Biol. 2020 Mar;102(4-5):537-551. doi: 10.1007/s11103-020-00963-7. Epub 2020 Jan 8. Plant Mol Biol. 2020. PMID: 31916084
-
Inflorescence development in female cannabis plants is mediated by photoperiod and gibberellin.Hortic Res. 2024 Sep 3;11(11):uhae245. doi: 10.1093/hr/uhae245. eCollection 2024 Nov. Hortic Res. 2024. PMID: 39539415 Free PMC article.
-
Transcriptome Sequencing-Based Analysis of Premature Fruiting in Amomum villosum Lour.Biology (Basel). 2025 Jul 18;14(7):883. doi: 10.3390/biology14070883. Biology (Basel). 2025. PMID: 40723439 Free PMC article.
-
Photoperiodic regulation of the sucrose transporter StSUT4 affects the expression of circadian-regulated genes and ethylene production.Front Plant Sci. 2013 Feb 20;4:26. doi: 10.3389/fpls.2013.00026. eCollection 2013. Front Plant Sci. 2013. PMID: 23429841 Free PMC article.
References
-
- Bagnall DJ. Control of flowering in Arabidopsis thaliana by light, vernalisation and gibberellins. Aust J Plant Physiol. 1992;19:401–409.
-
- Barber HN, Jackson WO, Murfet IC, Sprent JL. Gibberellic acid and the physiological genetics of flowering in peas. Nature. 1958;182:1321–1322.
-
- Bodson M. Changes in the carbohydrate content of the leaf and the apical bud of Sinapis during transition to flowering. Planta. 1977;135:19–23. - PubMed
-
- Bodson M, King RW, Evans LT, Bernier G. The role of photosynthesis in flowering of the long-day plant Sinapis alba. Aust J Plant Physiol. 1977;4:467–478.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
