

Excessive activation of serotonin (5-HT) 1B receptors disrupts the formation of sensory maps in monoamine oxidase a and 5-ht transporter knock-out mice
- PMID: 11157075
- PMCID: PMC6762299
- DOI: 10.1523/JNEUROSCI.21-03-00884.2001
Excessive activation of serotonin (5-HT) 1B receptors disrupts the formation of sensory maps in monoamine oxidase a and 5-ht transporter knock-out mice
Abstract
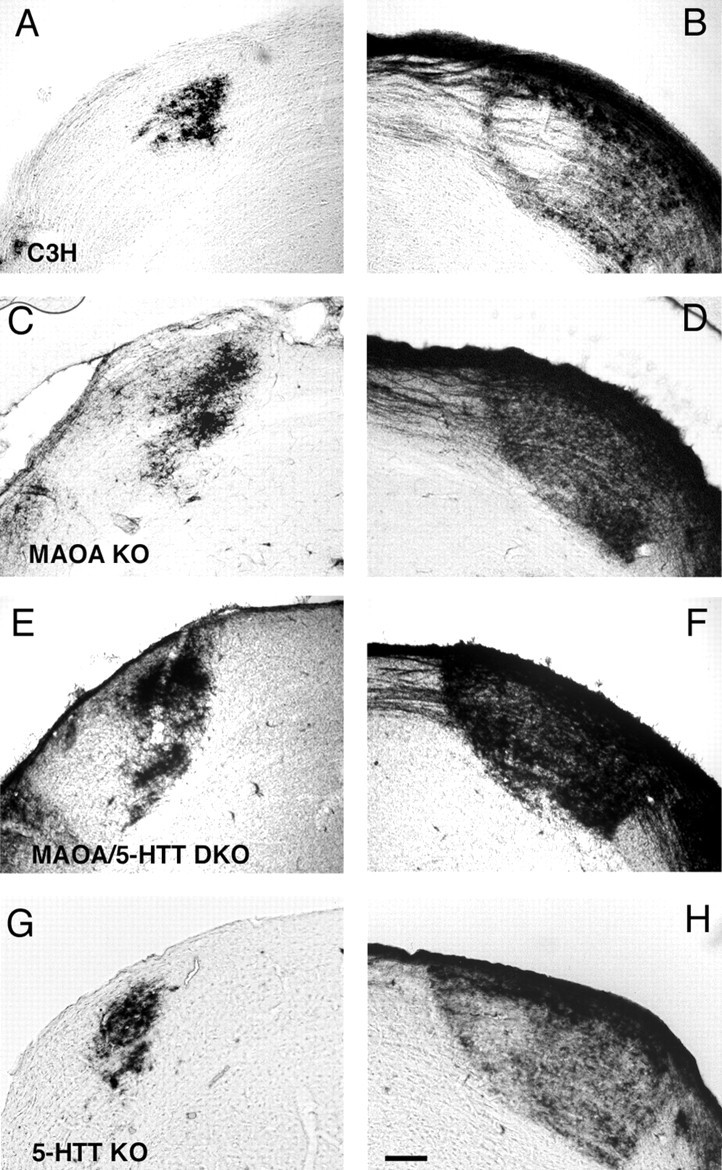
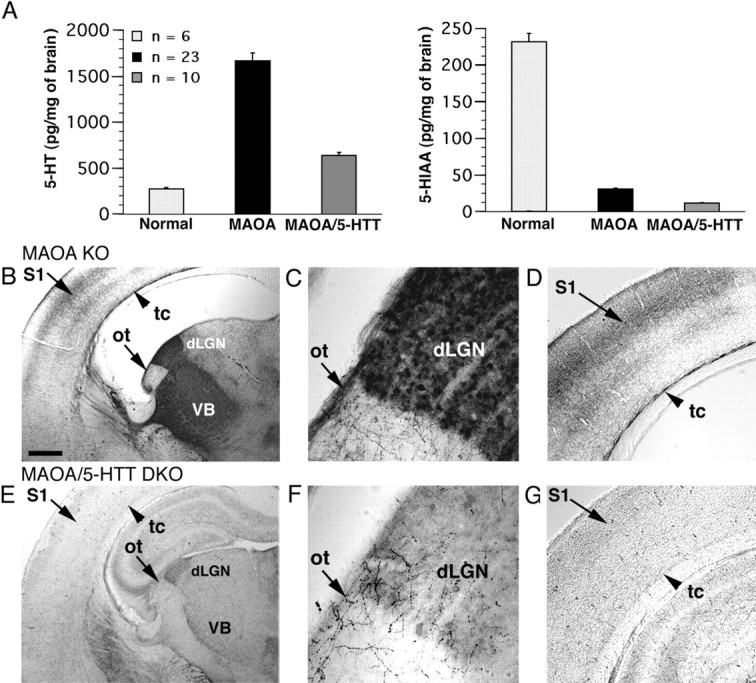
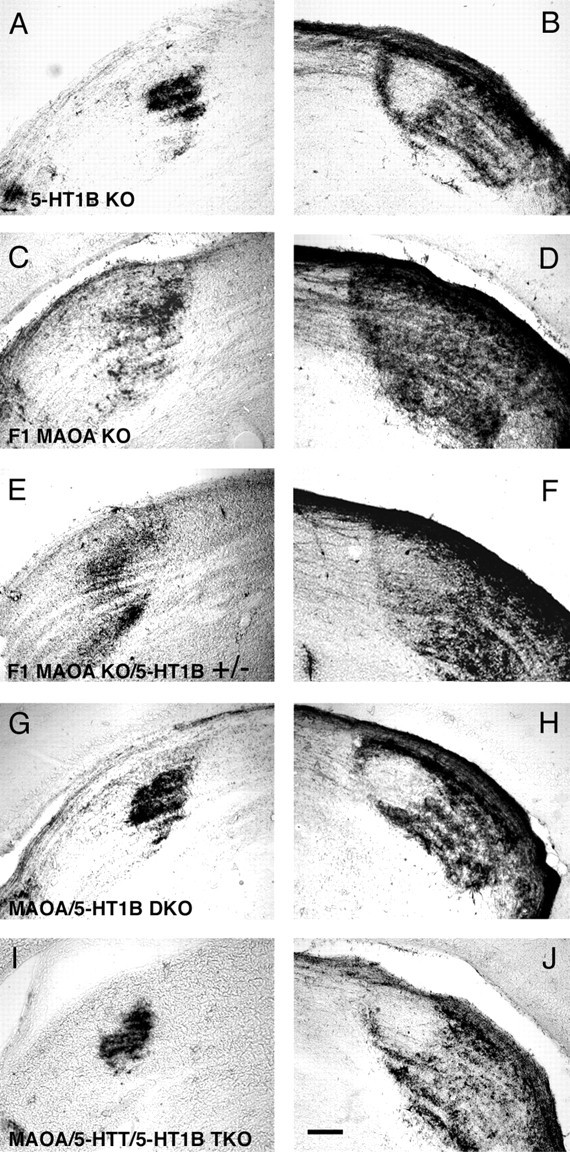
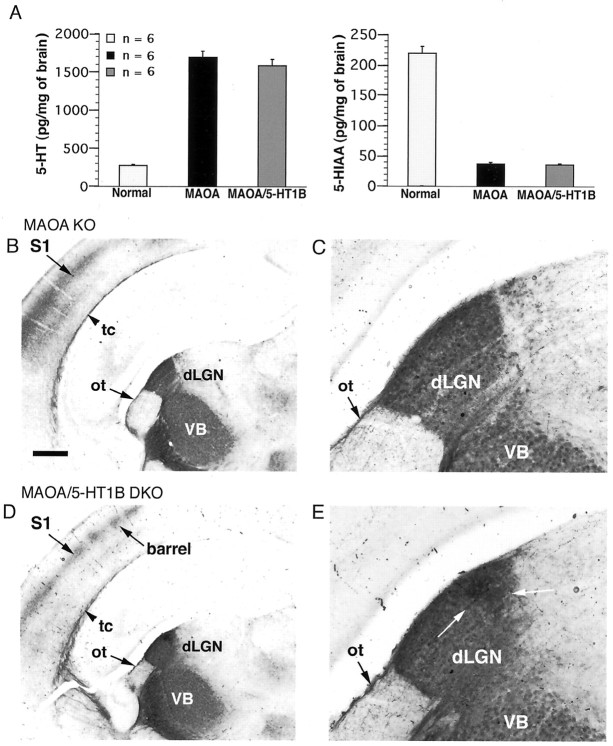
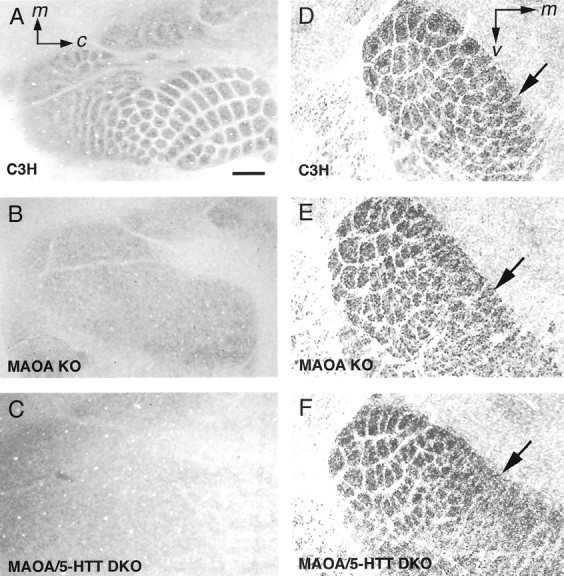
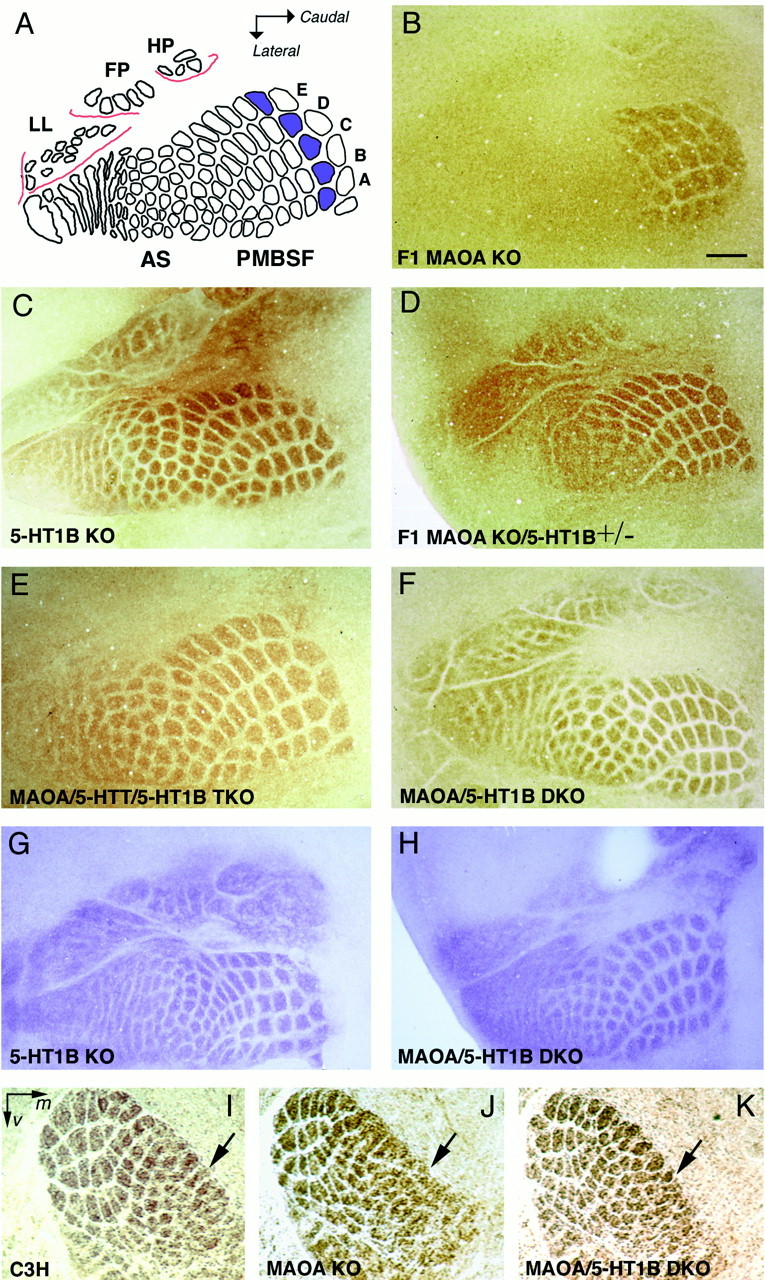
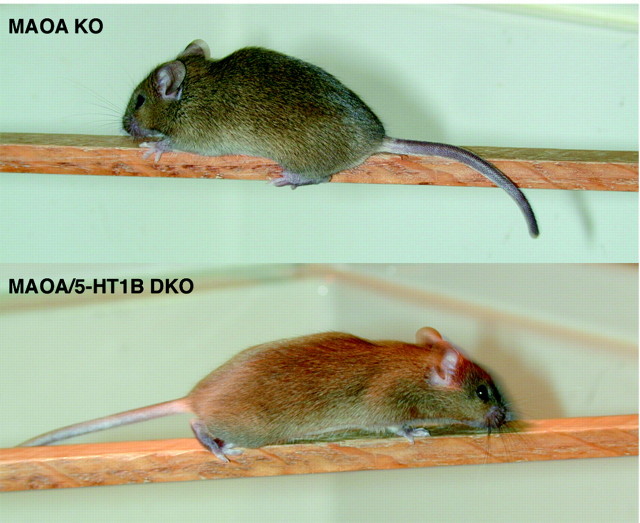
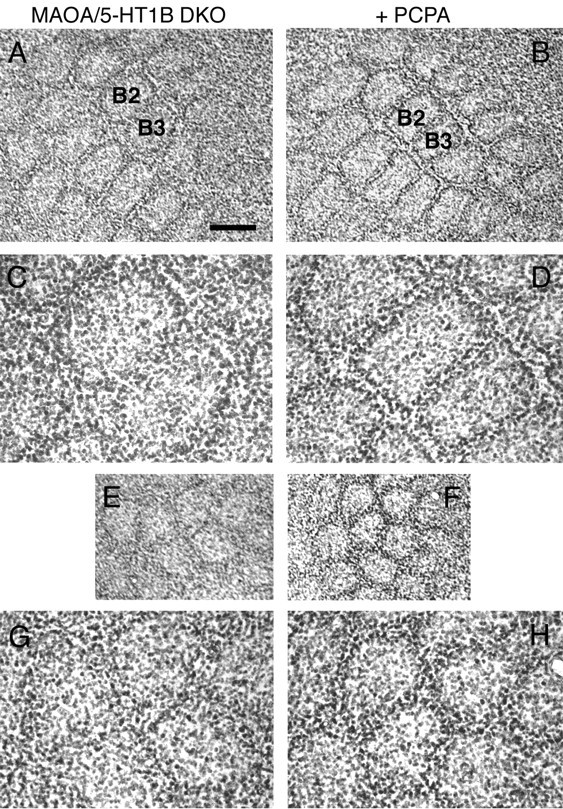
Deficiency in the monoamine degradation enzyme monoamine oxidase A (MAOA) or prenatal exposure to the monoamine uptake inhibitor cocaine alters behavior in humans and rodents, but the mechanisms are unclear. In MAOA knock-out mice, inhibiting serotonin synthesis during development can prevent abnormal segregation of axons in the retinogeniculate and somatosensory thalamocortical systems. To investigate this effect, we crossed MAOA knock-outs with mice lacking the serotonin transporter 5-HTT or the 5-HT1B receptor, two molecules present in developing sensory projections. Segregation was abnormal in 5-HTT knock-outs and MAOA/5-HTT double knock-outs but was normalized in MAOA/5-HT1B double knock-outs and MAOA/5-HTT/5-HT1B triple knock-outs. This demonstrates that the 5-HT1B receptor is a key factor in abnormal segregation of sensory projections and suggests that serotonergic drugs represent a risk for the development of these projections. We also found that the 5-HT1B receptor has an adverse developmental impact on beam-walking behavior in MAOA knock-outs. Finally, because the 5-HT1B receptor inhibits glutamate release, our results suggest that visual and somatosensory projections must release glutamate for proper segregation.
Figures
Similar articles
-
Lack of 5-HT(1B) receptor and of serotonin transporter have different effects on the segregation of retinal axons in the lateral geniculate nucleus compared to the superior colliculus.Neuroscience. 2002;111(3):597-610. doi: 10.1016/s0306-4522(01)00602-9. Neuroscience. 2002. PMID: 12031347
-
Refinement of thalamocortical arbors and emergence of barrel domains in the primary somatosensory cortex: a study of normal and monoamine oxidase a knock-out mice.J Neurosci. 2002 Oct 1;22(19):8541-52. doi: 10.1523/JNEUROSCI.22-19-08541.2002. J Neurosci. 2002. PMID: 12351728 Free PMC article.
-
Excess of serotonin (5-HT) alters the segregation of ispilateral and contralateral retinal projections in monoamine oxidase A knock-out mice: possible role of 5-HT uptake in retinal ganglion cells during development.J Neurosci. 1999 Aug 15;19(16):7007-24. doi: 10.1523/JNEUROSCI.19-16-07007.1999. J Neurosci. 1999. PMID: 10436056 Free PMC article.
-
[Implication of serotonin in the control of vigilance states as revealed by knockout-mouse studies].J Soc Biol. 2004;198(1):30-6. J Soc Biol. 2004. PMID: 15146953 Review. French.
-
Genetics of the serotonergic system in suicidal behavior.J Psychiatr Res. 2003 Sep-Oct;37(5):375-86. doi: 10.1016/s0022-3956(03)00048-7. J Psychiatr Res. 2003. PMID: 12849930 Review.
Cited by
-
Cortical RORβ is required for layer 4 transcriptional identity and barrel integrity.Elife. 2020 Aug 27;9:e52370. doi: 10.7554/eLife.52370. Elife. 2020. PMID: 32851975 Free PMC article.
-
Serotonin induces tonic firing in layer V pyramidal neurons of rat prefrontal cortex during postnatal development.J Neurosci. 2003 Apr 15;23(8):3373-84. doi: 10.1523/JNEUROSCI.23-08-03373.2003. J Neurosci. 2003. PMID: 12716945 Free PMC article.
-
Serotonin, genetic variability, behaviour, and psychiatric disorders--a review.Ups J Med Sci. 2010 Feb;115(1):2-10. doi: 10.3109/03009730903573246. Ups J Med Sci. 2010. PMID: 20187845 Free PMC article. Review.
-
Gene-environment interaction and the genetics of depression.J Psychiatry Neurosci. 2004 May;29(3):174-84. J Psychiatry Neurosci. 2004. PMID: 15173894 Free PMC article. Review.
-
Developmental role of tryptophan hydroxylase in the nervous system.Mol Neurobiol. 2007 Feb;35(1):45-54. doi: 10.1007/BF02700623. Mol Neurobiol. 2007. PMID: 17519505 Review.
References
-
- Adams JC. Stabilizing and rapid thionin staining of TMB-based HRP reaction product. Neurosci Lett. 1980;17:7–9. - PubMed
-
- Bengel D, Murphy DL, Andrews AM, Wichems CH, Feltner D, Heils A, Mössner R, Westphal H, Lesch KP. Altered brain serotonin homeostasis and locomotor insensitivity to 3,4-methylenedioxymethamp-hetamine (“Ecstasy”) in serotonin transporter-deficient mice. Mol Pharmacol. 1998;53:649–655. - PubMed
-
- Bennett-Clarke CA, Chiaia NL, Rhoades RW. Thalamocortical afferents in rat transiently express high-affinity serotonin uptake sites. Brain Res. 1996;733:301–306. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases