

Influenza B virus NS1 protein inhibits conjugation of the interferon (IFN)-induced ubiquitin-like ISG15 protein
- PMID: 11157743
- PMCID: PMC133459
- DOI: 10.1093/emboj/20.3.362
Influenza B virus NS1 protein inhibits conjugation of the interferon (IFN)-induced ubiquitin-like ISG15 protein
Abstract
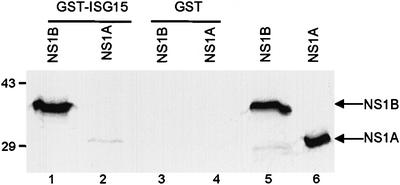
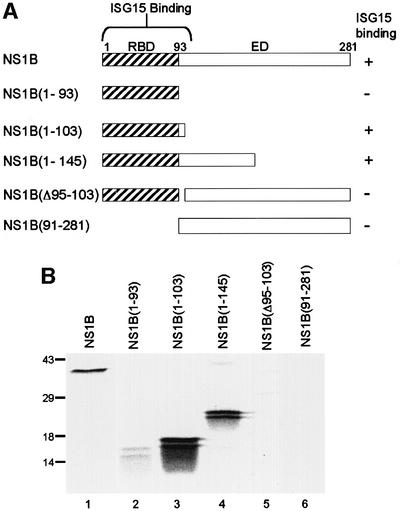
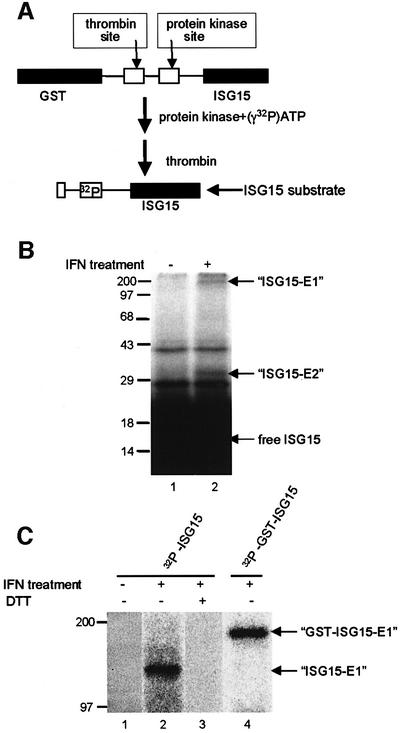
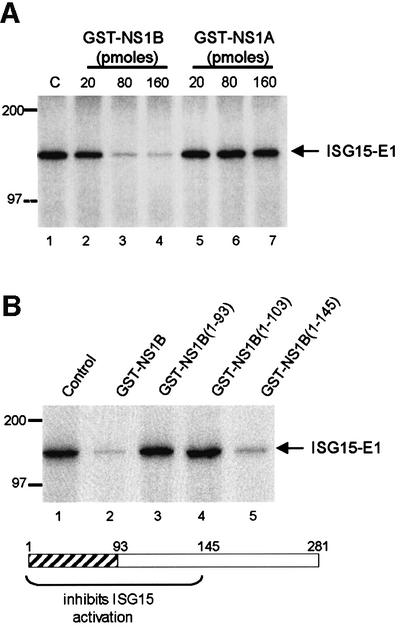
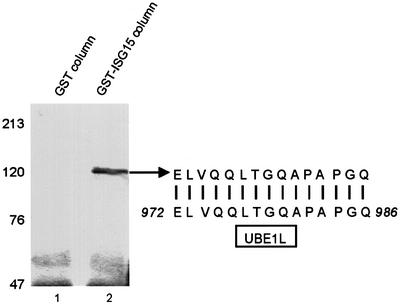
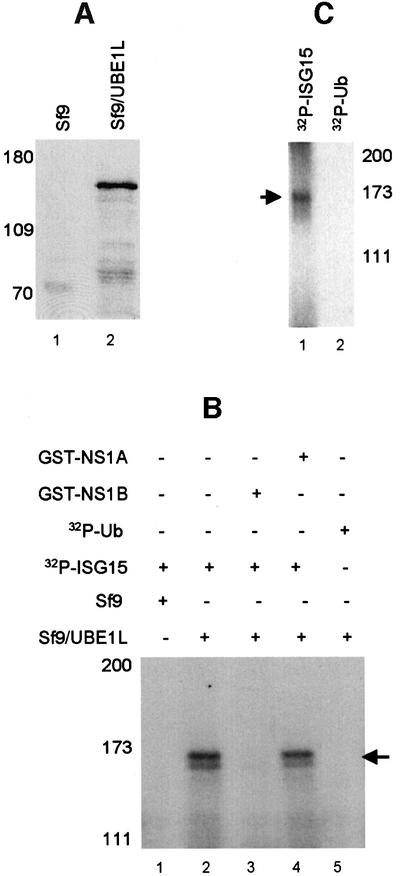
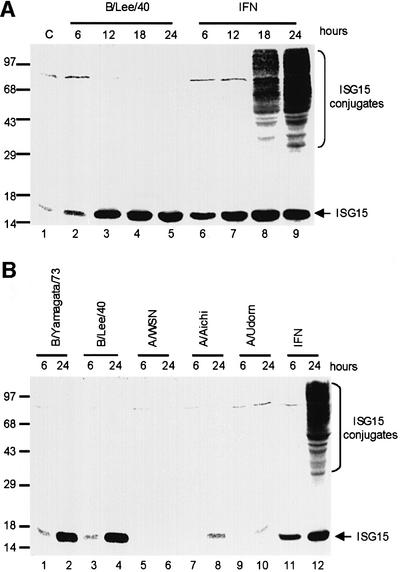
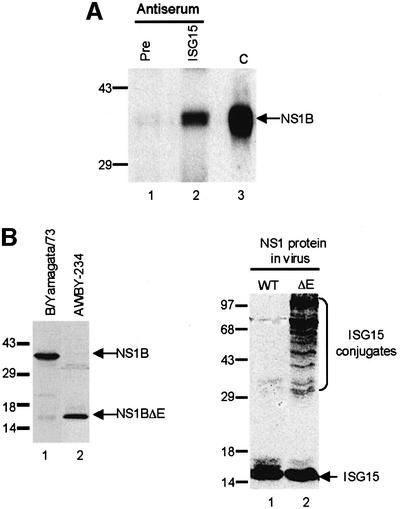
Of the several hundred proteins induced by interferon (IFN) alpha/beta, the ubiquitin-like ISG15 protein is one of the most predominant. We demonstrate the novel way in which the function of the ISG15 protein is inhibited by influenza B virus, which strongly induces the ISG15 protein: a specific region of the influenza B virus NS1 protein, which includes part of its effector domain, blocks the covalent linkage of ISG15 to its target proteins both in vitro and in infected cells. We identify UBE1L as the E1 enzyme that catalyzes the first activation step in the conjugation of ISG15, and show that the NS1B protein inhibits this activation step in vitro. Influenza A virus employs a different strategy: its NS1 protein does not bind the ISG15 protein, but little or no ISG15 protein is produced during infection. We discuss the likely basis for these different strategies.
Figures
References
-
- Beattie E.J., Tartaglia,J. and Paoletti,E. (1991) Vaccinia virus-encoded eIF-2α homolog abrogates the antiviral effects of interferon. Virology, 183, 419–422. - PubMed
-
- Carritt B. et al. (1992) A gene from human chromosome region 3p21 with reduced expression in small cell lung cancer. Cancer Res., 52, 1536–1541. - PubMed
-
- Chen Z. and Krug,R.M. (2000) Selective nuclear export of viral mRNAs in influenza-virus-infected cells. Trends Microbiol., 8, 376–383. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
