

Structural analysis of adenylate cyclases from Trypanosoma brucei in their monomeric state
- PMID: 11157750
- PMCID: PMC133460
- DOI: 10.1093/emboj/20.3.433
Structural analysis of adenylate cyclases from Trypanosoma brucei in their monomeric state
Erratum in
- EMBO J 2001 Sep 17;20(18):5302
Abstract
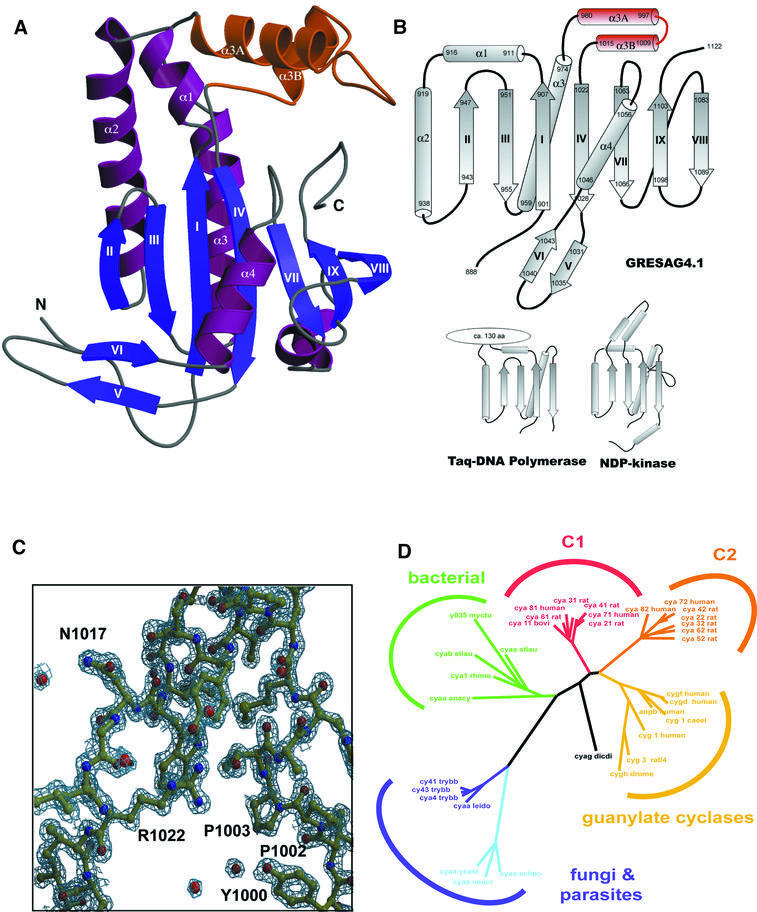
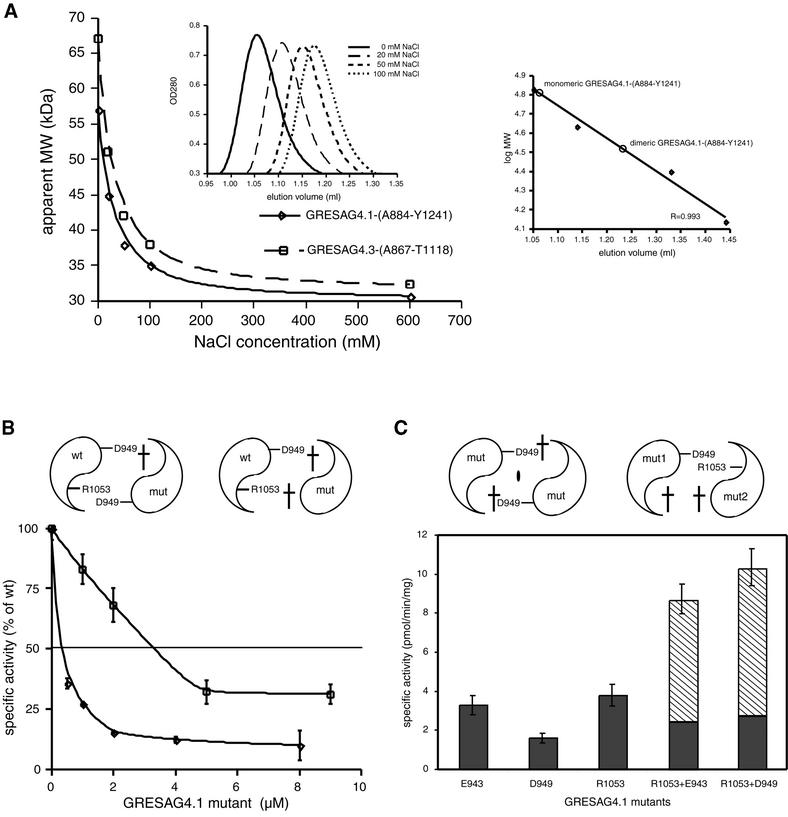
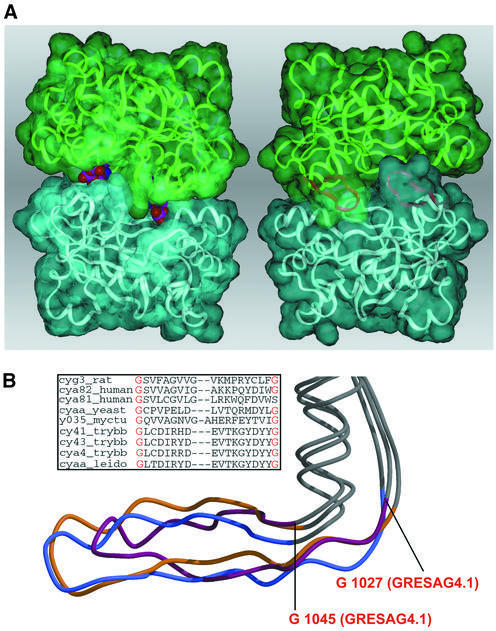
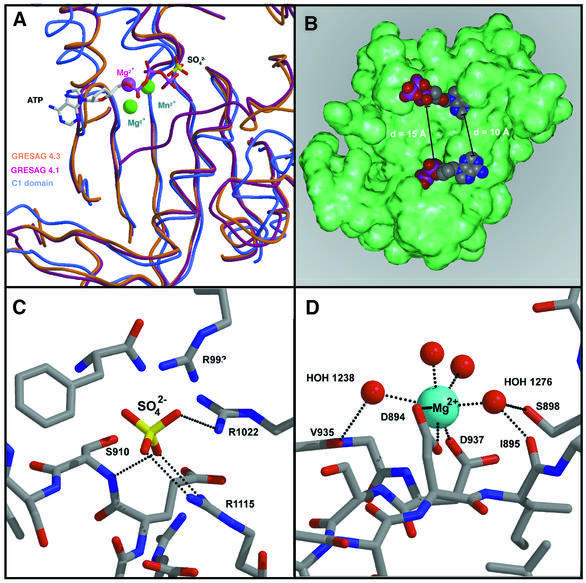
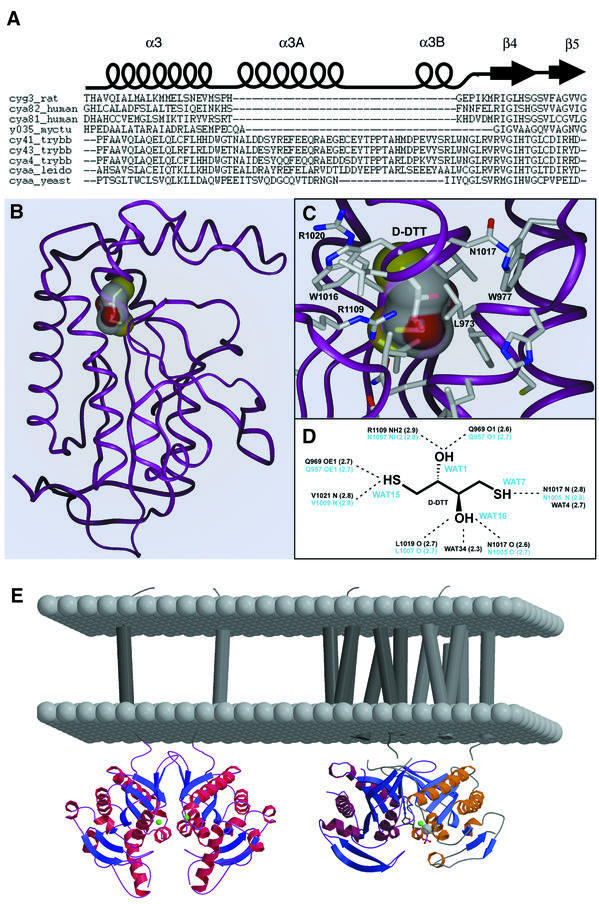
Cyclic AMP is a major trigger of the differentiation process of Trypanosoma brucei, a bloodstream parasite causing sleeping sickness. Its generation in trypanosomes is accomplished by a unique battery of membrane-bound adenylate cyclases (ACs). We have determined the high-resolution X-ray structures of the catalytic domains of two trypanosomal ACs (tACs), GRESAG4.1 and GRESAG4.3. The tAC domains are structurally highly related to the AC domains of higher eukaryotes, but also comprise a highly conserved structural element near the active site, the Delta-subdomain. A cavity below the Delta-subdomain might correspond to an allosteric regulator site as indicated by the stereospecific binding of a single (2S,3S)-1,4- dimercapto-2,3-butanediol molecule. In three different crystal forms, the tAC domains are exclusively observed in a monomeric, catalytically inactive state. Biochemical analysis and the mutagenesis profile of GRESAG4.1 confirmed a common catalytic mechanism of tACs that involves transient dimerization of the AC domain. A low dimerization tendency might play a regulatory role in T. brucei if the activation of tACs is similarly driven by ligand-induced dimerization as in membrane-bound guanylate cyclases.
Figures
References
-
- Abrahams J.P. and Leslie,A.G.W. (1996) Methods used in the structure determination of bovine mitochondrial F1-ATPase. Acta Crystallogr. D, 52, 30–42. - PubMed
-
- Alexandre S., Paindavoine,P., Tebabi,P., Pays,A., Halleux,S., Steinert,M. and Pays,E. (1990) Differential expression of a family of putative adenylate/guanylate cyclase genes in Trypanosoma brucei. Mol. Biochem. Parasitol., 43, 279–288. - PubMed
-
- Artymiuk P.J., Poirrette,A.R., Rice,D.W. and Willett,P. (1997) A polymerase I palm in adenylyl cyclase? Nature, 388, 33–34. - PubMed
-
- Barzu O. and Danchin,A. (1994) Adenylyl cyclases: a heterogeneous class of ATP-utilizing enzymes. Prog. Nucleic Acid Res. Mol. Biol., 49, 241–283. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
