

Genetic evidence for a role for Src family kinases in TNF family receptor signaling and cell survival
- PMID: 11157779
- PMCID: PMC312612
- DOI: 10.1101/gad.840301
Genetic evidence for a role for Src family kinases in TNF family receptor signaling and cell survival
Abstract
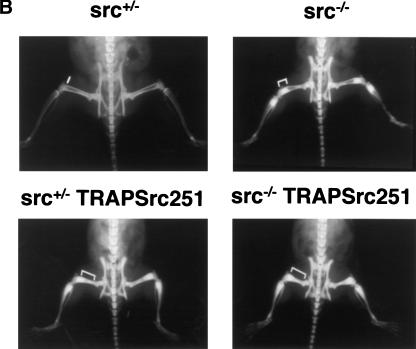
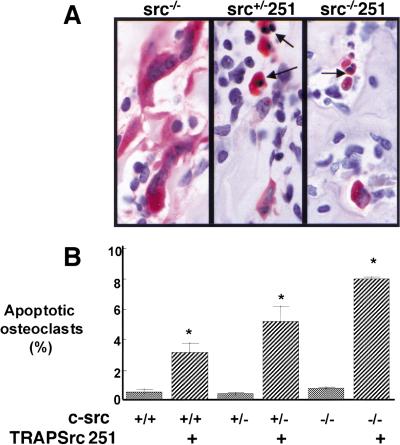
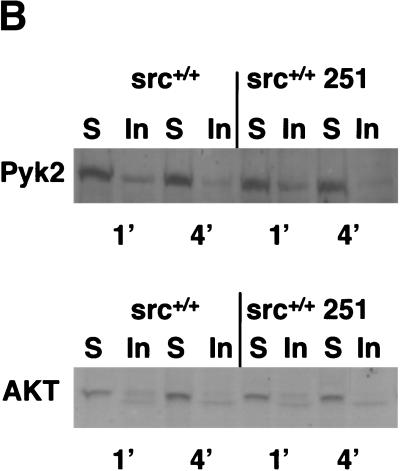
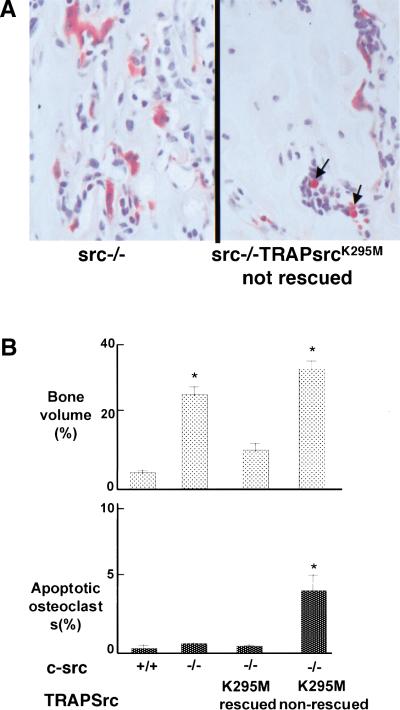
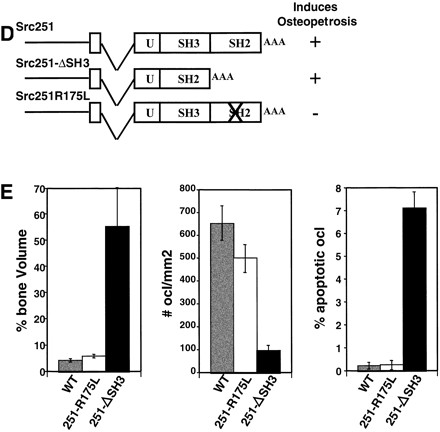
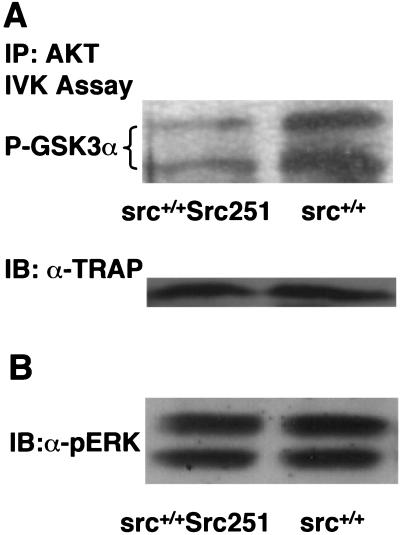
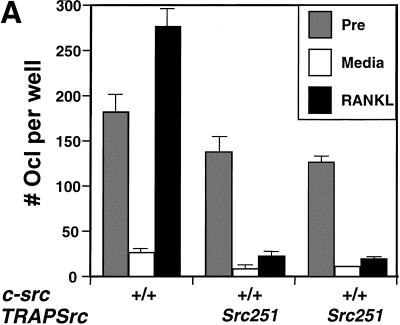
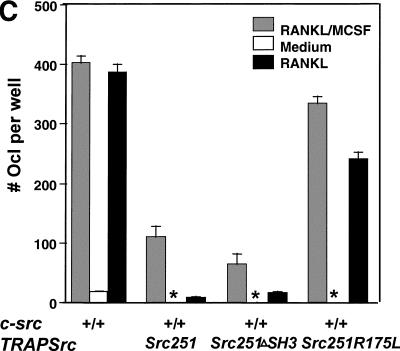
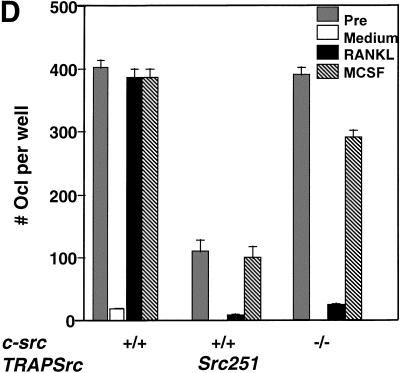
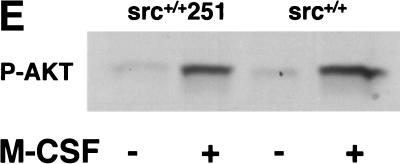
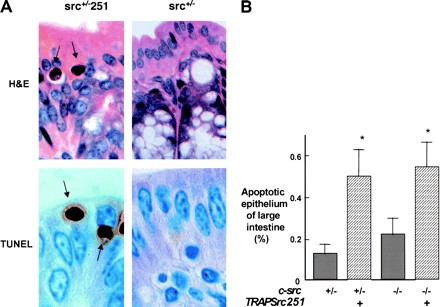
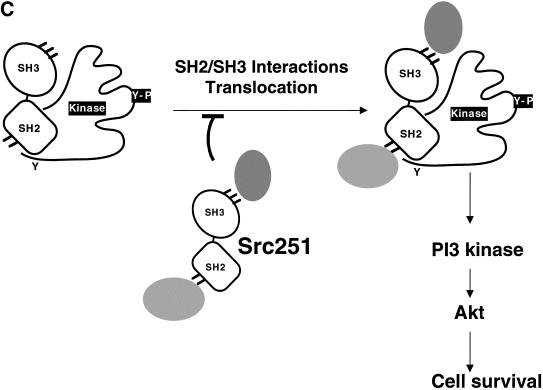
Mutant src(-/-) mice have osteopetrosis resulting from defective osteoclasts, the cells that resorb bone. However, signaling pathways involving Src family members in osteoclasts remain unclear. We demonstrate that expression of a truncated Src molecule, Src251, lacking the kinase domain, induces osteopetrosis in wild-type and src(+/-) mice and worsens osteopetrosis in src(-/-) mice by a novel mechanism, increased osteoclast apoptosis. Induction of apoptosis by Src251 requires a functional SH2, but not an SH3, domain and is associated with reduced AKT kinase activity. Expression of Src251 dramatically reduces osteoclast survival in response to RANKL/TRANCE/OPGL, providing evidence that Src family kinases are required in vivo for survival signaling pathways downstream from TNF family receptors.
Figures





References
-
- Brown MT, Cooper JA. Regulation, substrates, and functions of Src. Biochim Biophys Acta. 1996;1287:121–149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous