

Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilities
- PMID: 11157931
- PMCID: PMC94992
- DOI: 10.1128/JB.183.4.1195-1204.2001
Initiation of biofilm formation by Pseudomonas aeruginosa 57RP correlates with emergence of hyperpiliated and highly adherent phenotypic variants deficient in swimming, swarming, and twitching motilities
Abstract
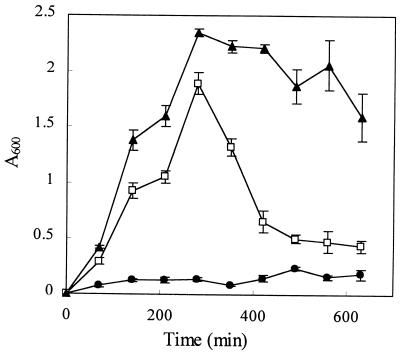
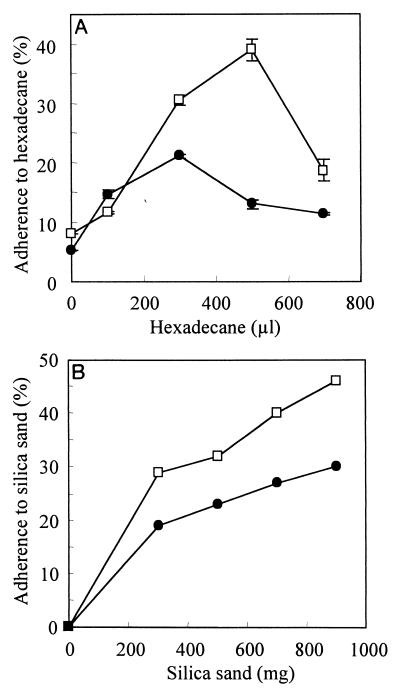
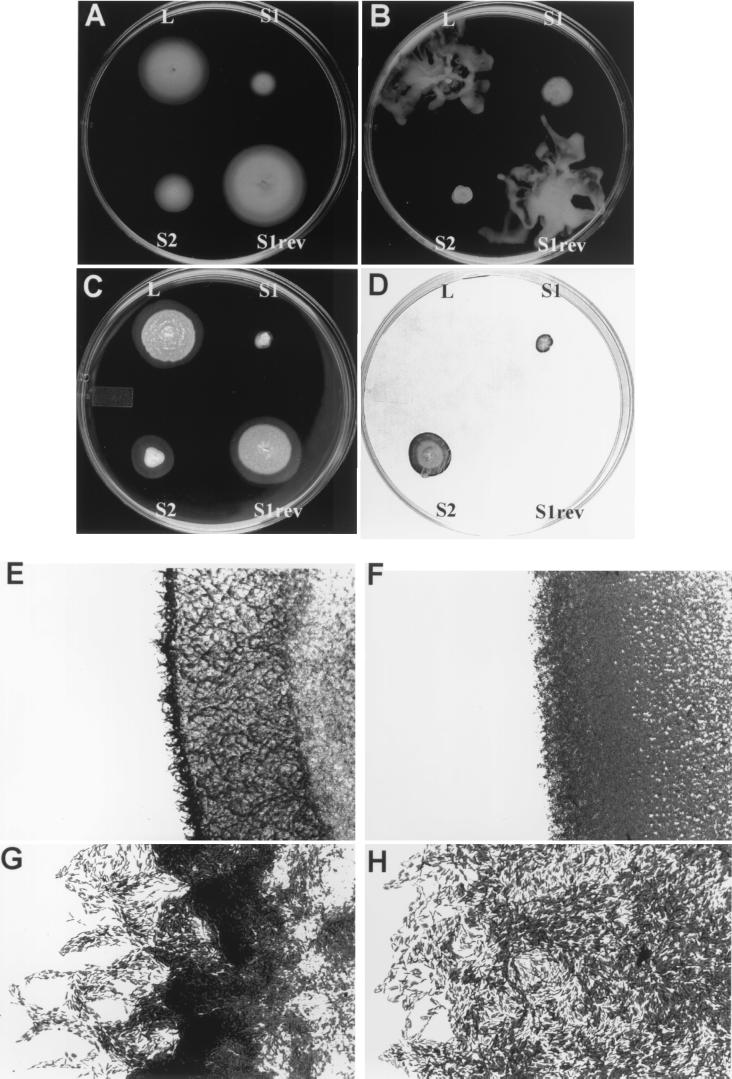
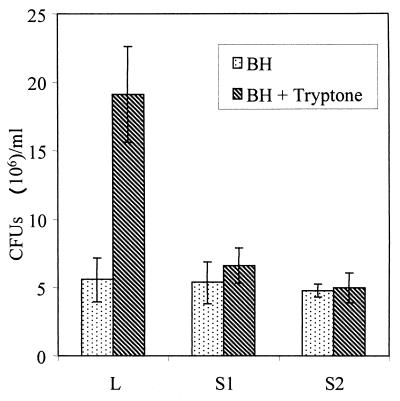
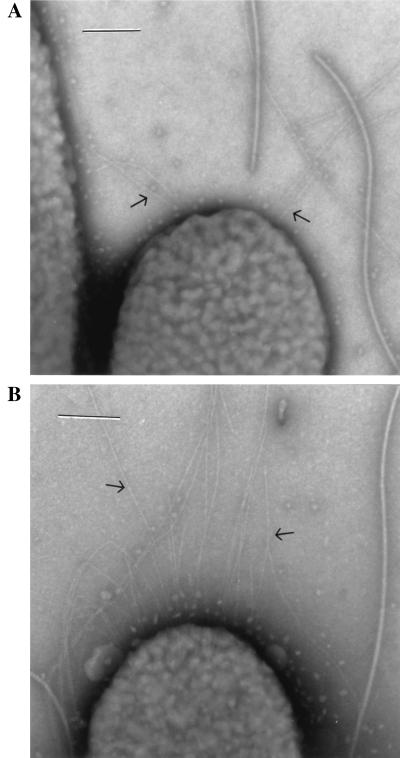
Pseudomonas aeruginosa is a ubiquitous environmental bacterium capable of forming biofilms on surfaces as a survival strategy. It exhibits a large variety of competition/virulence factors, such as three types of motilities: flagellum-mediated swimming, flagellum-mediated swarming, and type IV pilus-mediated twitching. A strategy frequently used by bacteria to survive changing environmental conditions is to create a phenotypically heterogeneous population by a mechanism called phase variation. In this report, we describe the characterization of phenotypic variants forming small, rough colonies that spontaneously emerged when P. aeruginosa 57RP was cultivated as a biofilm or in static liquid cultures. These small-colony (S) variants produced abundant type IV fimbriae, displayed defective swimming, swarming, and twitching motilities, and were impaired in chemotaxis. They also autoaggregated in liquid cultures and rapidly initiated the formation of strongly adherent biofilms. In contrast, the large-colony variant (parent form) was poorly adherent, homogeneously dispersed in liquid cultures, and produced scant polar fimbriae. Further analysis of the S variants demonstrated differences in a variety of other phenotypic traits, including increased production of pyocyanin and pyoverdine and reduced elastase activity. Under appropriate growth conditions, cells of each phenotype switched to the other phenotype at a fairly high frequency. We conclude that these S variants resulted from phase variation and were selectively enriched when P. aeruginosa 57RP was grown as a biofilm or in static liquid cultures. We propose that phase variation ensures the prior presence of phenotypic forms well adapted to initiate the formation of a biofilm as soon as environmental conditions are favorable.
Figures

References
-
- Alm R A, Mattick J S. Genes involved in the biogenesis and function of type-4 fimbriae in Pseudomonas aeruginosa. Gene. 1997;192:89–98. - PubMed
-
- Bradley D E. The adsorption of Pseudomonas aeruginosa pilus-dependent bacteriophages to a host mutant with nonretractile pili. Virology. 1974;58:149–163. - PubMed
-
- Chandrasekaran E V, BeMiller J N. Constituent analysis of glucosaminoglycans. In: Whistler R L, editor. Methods in carbohydrate chemistry. New York, N.Y: Academic Press, Inc.; 1980. pp. 89–96.
-
- Costerton J W, Lewandowski Z, Caldwell D E, Korber D R, Lappin-Scott H M. Microbial biofilms. Annu Rev Microbiol. 1995;49:711–745. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
