

Splicing factor slt11p and its involvement in formation of U2/U6 helix II in activation of the yeast spliceosome
- PMID: 11158289
- PMCID: PMC99556
- DOI: 10.1128/MCB.21.4.1011-1023.2001
Splicing factor slt11p and its involvement in formation of U2/U6 helix II in activation of the yeast spliceosome
Abstract
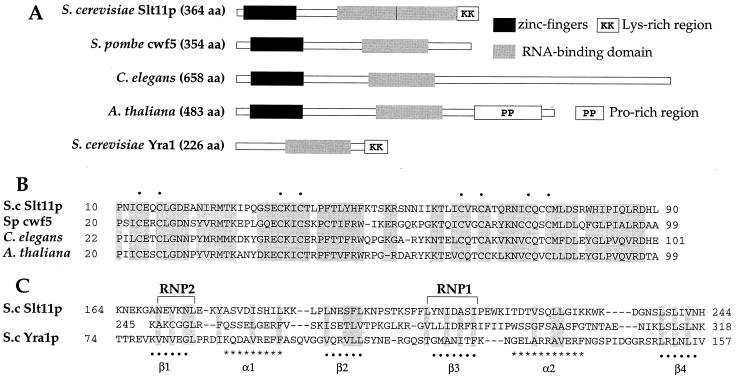
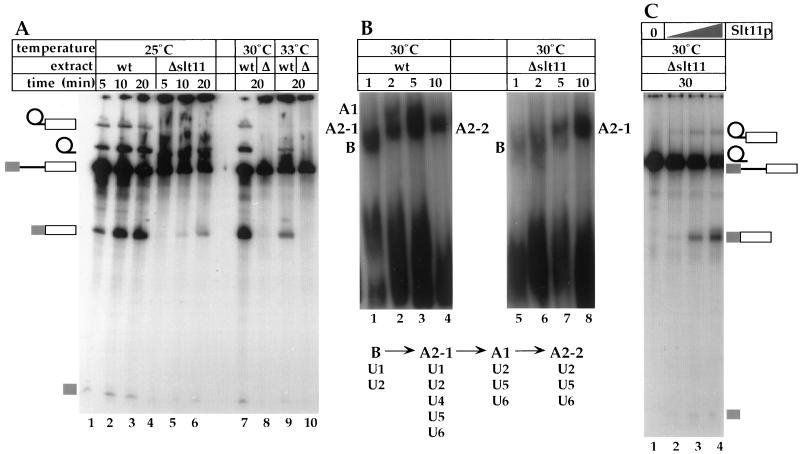
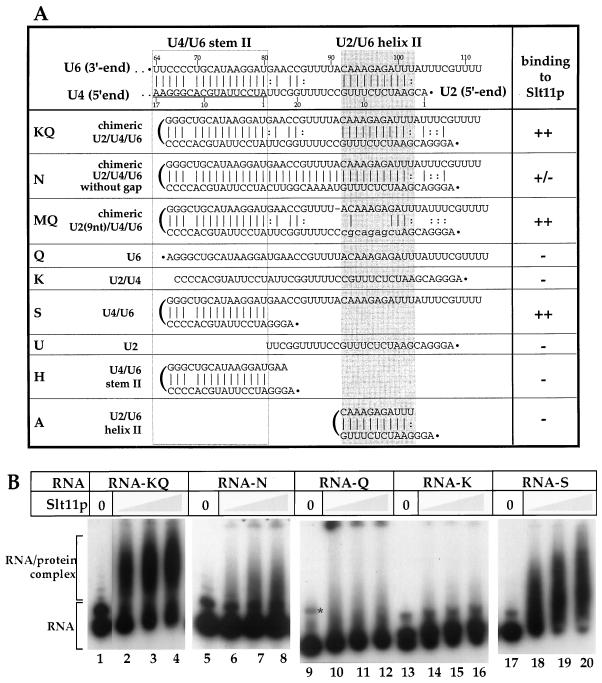
Slt11p is a new splicing factor identified on the basis of synthetic lethality with a mutation in the 5' end of U2 snRNA, a region that is involved in intermolecular U2/U6 helix II interaction. Slt11p is required for spliceosome assembly. Our genetic results suggest that Slt11p is involved in the base-pairing interaction of U2/U6 helix II in vivo. We showed that the recombinant protein binds to RNAs with some degree of structural specificity. Slt11p also anneals RNA and binds to the resulting duplexes, which contain two separated helical regions. These RNA structures are reminiscent of U2/U6 helix II, which is formed concomitantly with U4/U6 stem II, and suggest that Slt11p facilitates the cooperative formation of helix II in association with stem II in the spliceosome. We show that Slt11p and Slu7p, a second-step factor, interact with each other both in vivo and in vitro and that the binding of Slu7p to Slt11p impairs the RNA-binding activity of the latter. These results suggest that the function of Slt11p is regulated by Slu7p in the spliceosome.
Figures






References
-
- Abovich N, Rosbash M. Cross-intron bridging interactions in the yeast commitment complex are conserved in mammals. Cell. 1997;89:403–412. - PubMed
-
- Abovich N, Liao X C, Rosbash M. The yeast MUD2 protein: an interaction with PRP11 defines a bridge between commitment complexes and U2 snRNP addition. Genes Dev. 1994;8:843–854. - PubMed
-
- Adams A, Gottschling D E, Kaiser C A, Stearns T. Methods in yeast genetics. Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory Press; 1997.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases