

Candida albicans INT1-induced filamentation in Saccharomyces cerevisiae depends on Sla2p
- PMID: 11158313
- PMCID: PMC99580
- DOI: 10.1128/MCB.21.4.1272-1284.2001
Candida albicans INT1-induced filamentation in Saccharomyces cerevisiae depends on Sla2p
Abstract
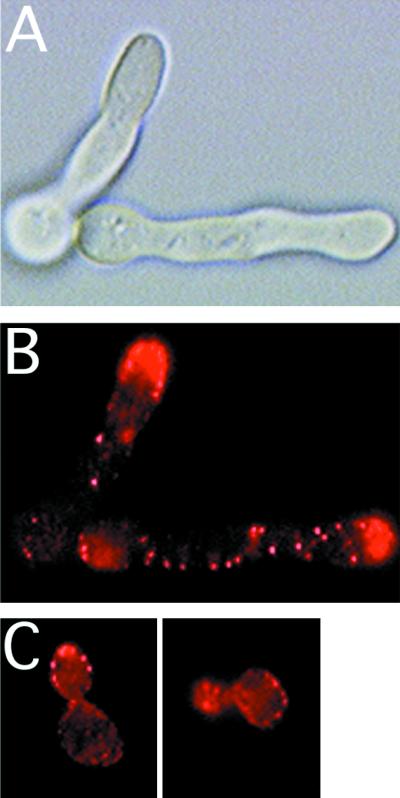
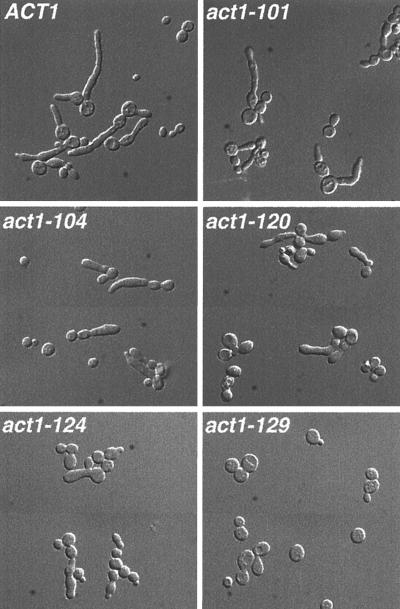
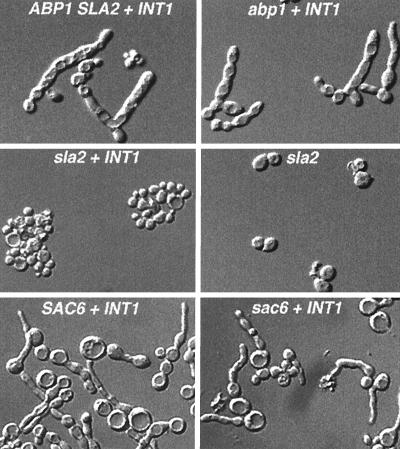
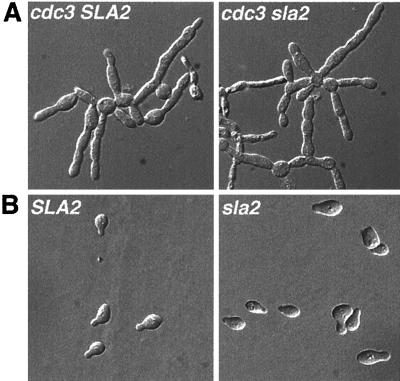
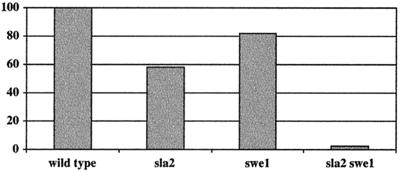
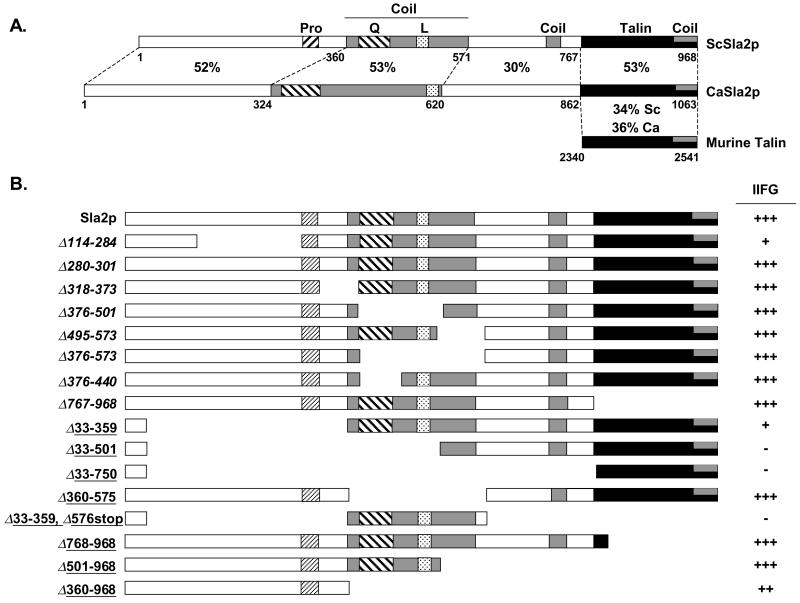
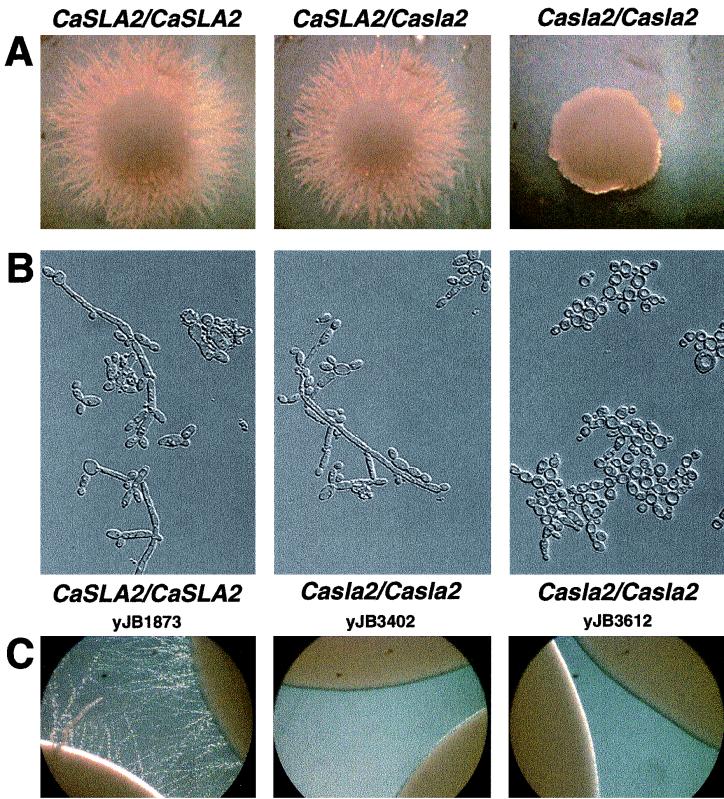
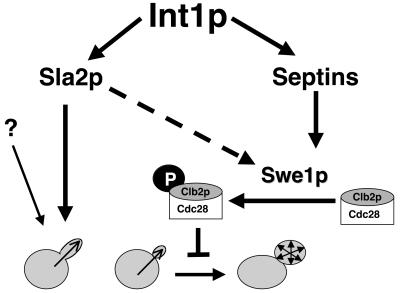
The Candida albicans INT1 gene is important for hyphal morphogenesis, adherence, and virulence (C. Gale, C. Bendel, M. McClellan, M. Hauser, J. M. Becker, J. Berman, and M. Hostetter, Science 279:1355-1358, 1998). The ability to switch between yeast and hyphal morphologies is an important virulence factor in this fungal pathogen. When INT1 is expressed in Saccharomyces cerevisiae, cells grow with a filamentous morphology that we exploited to gain insights into how C. albicans regulates hyphal growth. In S. cerevisiae, INT1-induced filamentous growth was affected by a small subset of actin mutations and a limited set of actin-interacting proteins including Sla2p, an S. cerevisiae protein with similarity in its C terminus to mouse talin. Interestingly, while SLA2 was required for INT1-induced filamentous growth, it was not required for polarized growth in response to several other conditions, suggesting that Sla2p is not required for polarized growth per se. The morphogenesis checkpoint, mediated by Swe1p, contributes to INT1-induced filamentous growth; however, epistasis analysis suggests that Sla2p and Swe1p contribute to INT1-induced filamentous growth through independent pathways. The C. albicans SLA2 homolog (CaSLA2) complements S. cerevisiae sla2Delta mutants for growth at 37 degrees C and INT1-induced filamentous growth. Furthermore, in a C. albicans Casla2/Casla2 strain, hyphal growth did not occur in response to either nutrient deprivation or to potent stimuli, such as mammalian serum. Thus, through analysis of INT1-induced filamentous growth in S. cerevisiae, we have identified a C. albicans gene, SLA2, that is required for hyphal growth in C. albicans.
Figures
References
-
- Adams A E, Pringle J R. Staining of actin with fluorochrome-conjugated phalloidin. Methods Enzymol. 1991;194:729–731. - PubMed
-
- Amberg D C, Basart E, Botstein D. Defining protein interactions with yeast actin in vivo. Nat Struct Biol. 1995;2:28–35. - PubMed
-
- Anderson J M, Soll D R. Differences in actin localization during bud and hypha formation in the yeast Candida albicans. J Gen Microbiol. 1986;132:2035–2047. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases