

Distinctive features of Drosophila alternative splicing factor RS domain: implication for specific phosphorylation, shuttling, and splicing activation
- PMID: 11158320
- PMCID: PMC99587
- DOI: 10.1128/MCB.21.4.1345-1359.2001
Distinctive features of Drosophila alternative splicing factor RS domain: implication for specific phosphorylation, shuttling, and splicing activation
Abstract
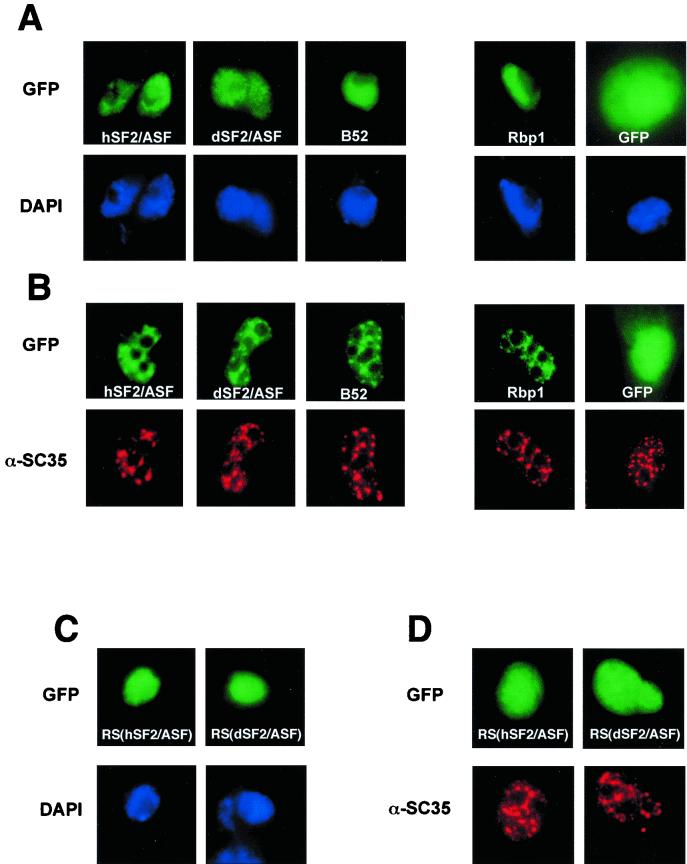
The human splicing factor 2, also called human alternative splicing factor (hASF), is the prototype of the highly conserved SR protein family involved in constitutive and regulated splicing of metazoan mRNA precursors. Here we report that the Drosophila homologue of hASF (dASF) lacks eight repeating arginine-serine dipeptides at its carboxyl-terminal region (RS domain), previously shown to be important for both localization and splicing activity of hASF. While this difference has no effect on dASF localization, it impedes its capacity to shuttle between the nucleus and cytoplasm and abolishes its phosphorylation by SR protein kinase 1 (SRPK1). dASF also has an altered splicing activity. While being competent for the regulation of 5' alternative splice site choice and activation of specific splicing enhancers, dASF fails to complement S100-cytoplasmic splicing-deficient extracts. Moreover, targeted overexpression of dASF in transgenic flies leads to higher deleterious developmental defects than hASF overexpression, supporting the notion that the distinctive structural features at the RS domain between the two proteins are likely to be functionally relevant in vivo.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials