

Extrafloral nectar production of the ant-associated plant, Macaranga tanarius, is an induced, indirect, defensive response elicited by jasmonic acid
- PMID: 11158598
- PMCID: PMC14712
- DOI: 10.1073/pnas.98.3.1083
Extrafloral nectar production of the ant-associated plant, Macaranga tanarius, is an induced, indirect, defensive response elicited by jasmonic acid
Abstract
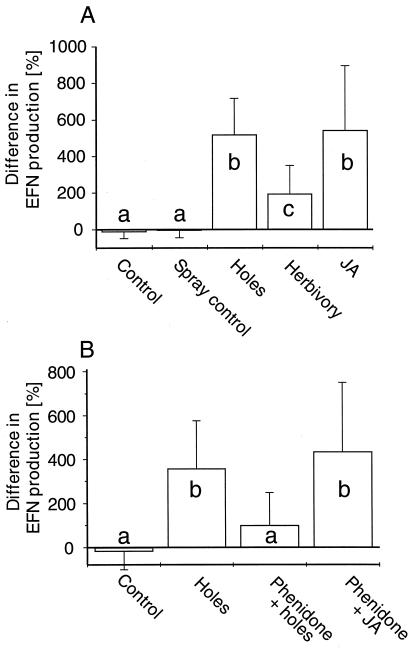
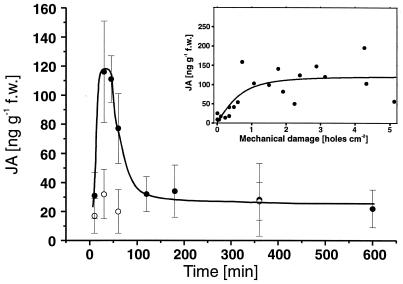
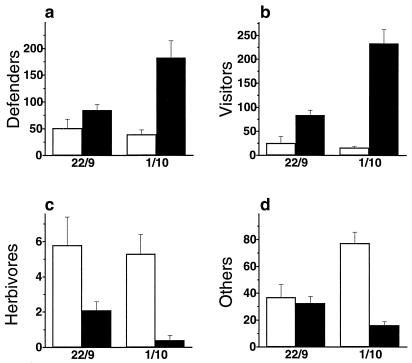
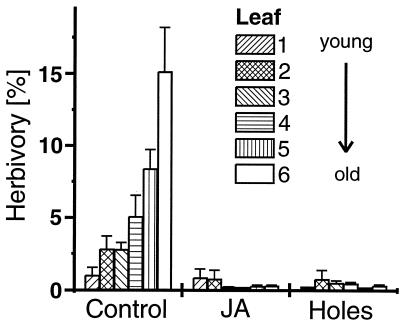
Plant species in at least 66 families produce extrafloral nectar (EFN) on their leaves or shoots and therewith attract predators and parasitoids, such as ants and wasps, which in turn defend them against herbivores. We investigated whether EFN secretion is induced by herbivory and/or artificial damage, and thus can be regarded as an induced defensive response. In addition, we studied the underlying signaling pathway. EFN secretion by field-grown Macaranga tanarius increased after herbivory, artificial leaf damage, and exogenous jasmonic acid (JA) application. Artificial damage strongly enhanced endogenous JA concentrations. The response in EFN production to artificial damage was much less pronounced in those leaves that were treated with phenidone to inhibit endogenous JA synthesis. Quantitative dose-response relations were found between the increase in nectar production and both the intensity of leaf damage and the amounts of exogenously applied JA. The amount of endogenously produced JA was positively correlated with the intensity of leaf damage. Increased numbers of defending insects and decreased numbers of herbivores were observed on leaves after inducing EFN production by exogenous JA treatment. Over 6 weeks, repeatedly applied JA or artificial damage resulted in a ten-fold reduction in herbivory. These results demonstrate that EFN production represents an alternative mechanism for induced, indirect defensive plant responses that are mediated via the octadecanoid signal transduction cascade.
Figures
Comment in
-
Polycystic kidney disease: In danger of being X-rated?Proc Natl Acad Sci U S A. 2001 Jan 30;98(3):790-2. doi: 10.1073/pnas.98.3.790. Proc Natl Acad Sci U S A. 2001. PMID: 11158545 Free PMC article. Review. No abstract available.
References
-
- Bentley B L. Ann Rev Ecol Syst. 1977;8:407–427.
-
- Elias T S. In: The Biology of Nectaries. Bentley B, Elias T S, editors. New York: Columbia Univ. Press; 1983. pp. 174–203.
-
- Buckley R C. In: Ant-Plant Interactions in Australia. Buckley R C, editor. The Hague, The Netherlands: W. Junk; 1982. pp. 111–162.
-
- Koptur S. In: Insect-Plant Interactions. Bernays E A, editor. Vol. 4. Boca Raton, FL: CRC Press; 1992. pp. 81–129.
-
- Barton A M. Ecology. 1986;67:495–504.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
