

Neural mechanisms underlying migrating motor complex formation in mouse isolated colon
- PMID: 11159701
- PMCID: PMC1572567
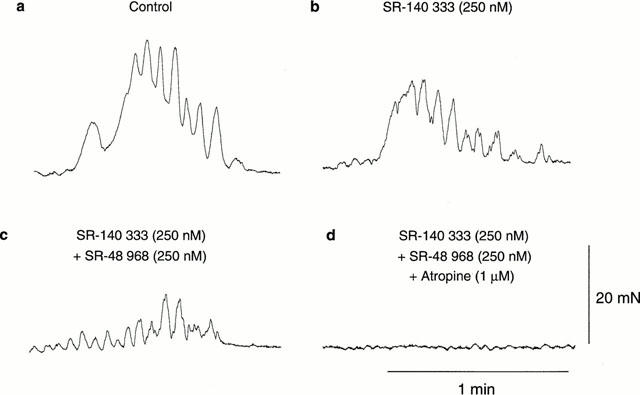
- DOI: 10.1038/sj.bjp.0703814
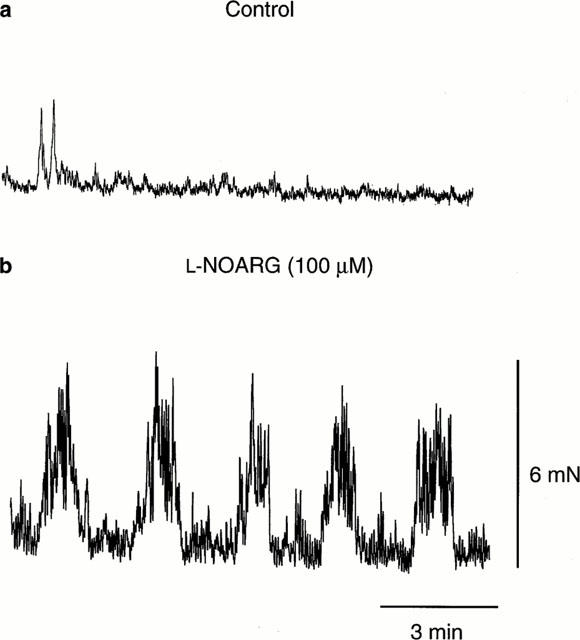
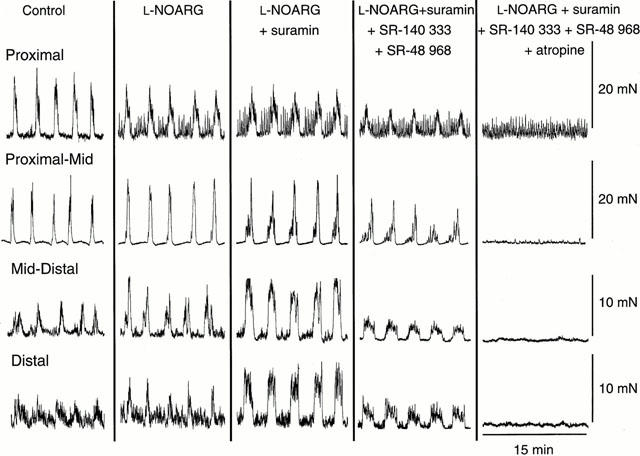
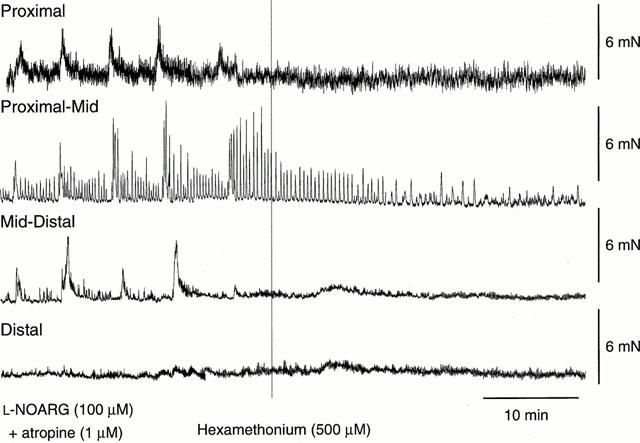
Neural mechanisms underlying migrating motor complex formation in mouse isolated colon
Abstract
1. Little is known about the intrinsic enteric reflex pathways associated with migrating motor complex (MMC) formation. Acetylcholine (ACh) mediates the rapid component of the MMC, however a non-cholinergic component also exists. The present study investigated the possible role of endogenous tachykinins (TKs) in the formation of colonic MMCs and the relative roles of excitatory and inhibitory pathways. 2. MMCs were recorded from the circular muscle at four sites (proximal, proximal-mid, mid-distal and distal) along the mouse colon using force transducers. 3. The tachykinin (NK(1) and NK(2)) receptor antagonists SR-140 333 (250 nM) and SR-48 968 (250 nM) reduced the amplitude of MMCs at all recording sites, preferentially abolishing the long duration contraction. Residual MMCs were abolished by the subsequent addition of atropine (1 microM). 4. The neuronal nitric oxide synthase inhibitor, N(omega)nitro-L-arginine (L-NOARG, 100 microM), increased MMC amplitude in the distal region, whilst reducing the amplitude in the proximal region. In preparations where MMCs did not migrate to the distal colon, addition of L-NOARG resulted in the formation of MMCs. Subsequent addition of apamin (250 nM) or suramin (100 microM) further increased MMC amplitude in the distal region, whilst suramin increased MMC amplitude in the mid-distal region. Apamin but not suramin reduced MMC amplitude in the proximal region. Subsequent addition of SR-140 333 and SR-48 968 reduced MMC amplitude at all sites. Residual MMCs were abolished by atropine (1 microM). 5. In conclusion, TKs, ACh, nitric oxide (NO) and ATP are involved in the neural mechanisms underlying the formation of MMCs in the mouse colon. Tachykinins mediate the long duration component of the MMC via NK(1) and NK(2) receptors. Inhibitory pathways may be involved in determining whether MMCs are formed.
Figures




References
-
- BARAJAS-LÓPEZ C., MULLER M.J., PRIETO-GÓMEZ B., ESPINOSA-LUNA R. ATP inhibits the synaptic release of acetylcholine in submucosal neurons. J. Pharmacol. Exp. Ther. 1995;274:1238–1245. - PubMed
-
- CLARK S.R., COSTA M., TONINI M., BROOKES S.J.H. Purinergic transmission is involved in a descending excitatory reflex in the guinea-pig ileum. Proc. Aust. Neurosci. Soc. 1996;7:176.
-
- COULIE B., CAMILLERI M. Intestinal pseudo-obstruction. Annu. Rev. Med. 1999;50:37–55. - PubMed
-
- CROCI T., EMONDS-ALT X., LEFUR G., MANARA L. In vitro characterization of the non-peptide tachykinin NK1 and NK2-receptor antagonists, SR140333 and SR48968 in different rat and guinea-pig intestinal segments. Life Sci. 1994;56:267–275. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
