

doi: 10.1101/gad.871501.
Wnt signals from the neural tube block ectopic cardiogenesis
Affiliations
- PMID: 11159906
- PMCID: PMC312627
- DOI: 10.1101/gad.871501
Item in Clipboard
Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev.
.
Abstract
It has long been observed that repressive signals from the neural tube block cardiogenesis in vertebrates. Here we show that a signal from the neural tube that blocks cardiogenesis in the adjacent anterior paraxial mesoderm of stage 8-9 chick embryos can be mimicked by ectopic expression of either Wnt-3a or Wnt-1, both of which are expressed in the dorsal neural tube. Repression of cardiogenesis by the neural tube can be overcome by ectopic expression of a secreted Wnt antagonist. On the basis of both in vitro and in vivo results, we propose that Wnt signals from the neural tube normally act to block cardiogenesis in the adjacent anterior paraxial mesendoderm.
Figures
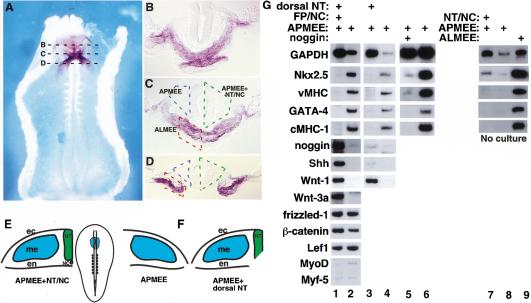
Signals from the dorsal neural tube block cardiogenesis in anterior paraxial mesendoderm. (A) Whole-mount in situ hybridization for Nkx-2.5 in a stage 9 chick embryo (ventral side up). (B–D) show representative transverse sections as indicated in A. Anterior paraxial mesendoderm with overlying ectoderm (APMEE) is outlined in blue, APMEE with adjacent neural tube and notochord is outlined in green, and the anterior lateral mesendoderm with overlying ectoderm (ALMEE) is outlined in red. (E) A diagram of a stage 9 chick embryo is shown in the middle panel, and the APMEE is indicated by blue shading. Diagrams of transverse sections through APMEE explants cultured in either the presence or the absence of the axial tissues are shown on the left and right, respectively. (F) Diagram of APMEE explant cultured in the presence of only the dorsal neural tube. (G) RT–PCR analysis of gene expression in explants of APMEE that have been cultured in vitro for 48 h in either the presence (lane 1) or the absence (lane 2) of the adjacent axial tissues or cultured in the presence (lane 3) or the absence (lane 4) of only the dorsal neural tube. Cardiogenesis was observed in 33 of 48 APMEE explants cultured in the absence of the axial tissues and was never observed in APMEE explants cultured in the presence of the axial tissues. APMEE explants were cultured in either the presence (lane 5) or absence (lane 6) of the BMP antagonist, noggin. Noggin administration blocked cardiogenesis in 10 out of 14 APMEE explants. Alternatively, explants of APMEE plus the neural tube and notochord (lane 7), APMEE alone (lane 8), or anterior lateral mesendoderm plus ectoderm (ALMEE; lane 9) were dissected and immediately harvested for RNA. Transcript levels of the indicated genes were monitored by RT–PCR analysis. Similar results were obtained in four independent experiments.
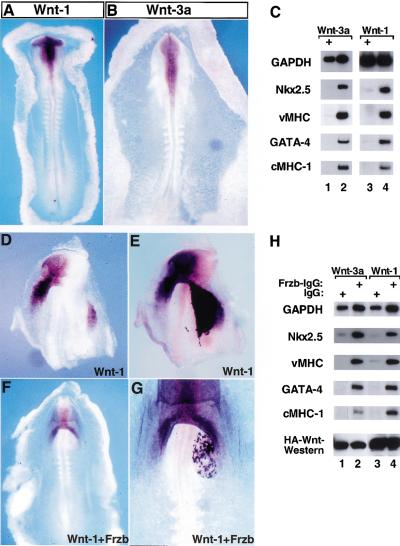
Inhibition of cardiogenesis by Wnt-1 and Wnt-3a. Expression of Wnt-1 (A) and Wnt-3a (B) as assessed by whole-mount in situ hybridization in stage 9–10 chick embryos. (C) Stage 9 APMEE explants were infected with either RCAS–Wnt-3a (lane 1) or RCAS–AP (lane 2). Rat-1/Wnt-1-HA cells (lane 3) or Rat-1 control cells (lane 4) were cocultured with APMEE explants on raft filters. Wnt-3a expression blocked cardiogenesis in ∼70% of the APMEE explants (n = 24), whereas Wnt-1 expression blocked cardiogenesis in 50% of such cultures (n = 12). (D–G) Rat-1/Wnt-1 cells were transiently transfected with either a control IgG expression vehicle (D,E) or a Frzb–IgG expression vehicle (F,G). Cell pellets were implanted into the left side of the heart-forming region of a stage 7 chick embryo that was maintained in New culture. Embryos were allowed to developed to stage 9–10, and subsequently analyzed by whole-mount in situ hybridization for Nkx-2.5 gene expression (D,F). Following in situ hybridization, the embryos were subsequently immunostained for IgG to identify the location of the IgG- or the Frzb–IgG-expressing cell pellet (E,G). The IgG- or the Frzb–IgG-expressing cells stain darker purple than cells expressing Nkx-2.5 as detected by in situ hybridization in D and F. (H) APMEE explants were cultured either with Rat-1/Wnt-3a-HA cells (lanes 1,2) or Rat-1/Wnt-1-HA cells (lanes 3,4) that had been transiently transfected with either control IgG (lanes 1,3) or with Frzb–IgG (lanes 2,4). After 48 h in culture, RNA was harvested and transcript levels of the indicated genes were monitored by RT–PCR analysis. Similar results were obtained in three independent experiments. Western blot analysis of the expression levels of the HA-tagged Wnts is shown.
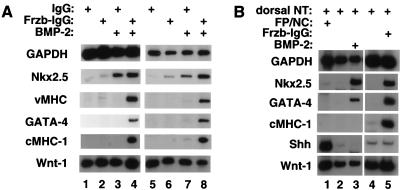
A combination of BMP signals and Frzb promotes cardiogenesis in anterior paraxial mesendoderm in the presence of the neural tube and notochord. (A) Stage 8 APMEE plus the adjacent neural tube and notochord (schematically illustrated in Fig. 1E) were dissected. Explants were cultured in the presence of either soluble control IgG (lanes 1,3) or soluble Frzb–IgG (lanes 2,4) in either the absence (lanes 1,2) or presence (lanes 3,4) of 60 ng/mL BMP-2. Alternatively, APMEE plus neural tube and notochord explants were cocultured on raft filters with aggregates of 293 cells transfected with either a control IgG expression vehicle (lanes 5,7) or a Frzb–IgG expression vehicle (lanes 6,8) and cultured either in the absence (lanes 5,6) or the presence (lanes 7,8) of 60 ng/mL BMP-2. After 48 h in culture, RNA was harvested and transcript levels monitored by RT–PCR. (B) Analysis of gene expression in APMEE explants that have been cultured in the presence of either the dorsal neural tube plus the floor plate and notochord (lane 1) or the dorsal neural tube only (lanes 2–5; schematically illustrated in Fig. 1F). Explants were cultured in the presence of exogenous BMP-2 or Frzb–IgG (lanes 3 and 5, respectively). Similar results have been obtained in four independent experiments.
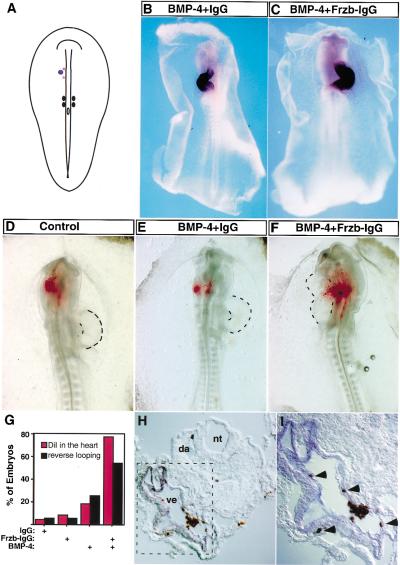
Inhibiting Wnt signals in vivo directs anterior paraxial mesodermal cells into the cardiac fate. (A) Pellets of cells expressing BMP-4, Frzb–IgG, and/or control IgG were implanted into the left side of the anterior paraxial mesoderm in stage 7 chick embryos. The location of the cell pellet is represented by a blue dot. In some cases, DiI was subsequently injected into a region that lay medial to the implanted cell pellet (red stars). The embryo is depicted dorsal side up. (B,C) Whole-mount in situ hybridization for vMHC expression is shown in stage 12 embryos that had been previously implanted with cell pellets expressing BMP-4 plus control IgG (B) or the combination of Frzb–IgG and BMP-4 (C). (D–F) Examples of the results obtained from DiI-injected embryos; dorsal side up. HEK-293 cells transfected with the indicated plasmids were implanted as described in A. Embryos were allowed to developed to stage 12–13 (26–30 h) before fixation. Brightfield and fluorescent images were taken and are overlayed. (G) Statistical summary of the in vivo results (red, DiI tracing; black, heart looping). (H,I) Implanting Frzb–IgG- and BMP-4-expressing cells into the anterior paraxial mesoderm induces migration of cells within this tissue into regions of the heart that express vMHC. Transverse section of embryo implanted with Frzb–IgG- and BMP-4-expressing cells in the anterior paraxial mesoderm (as shown in F). DiI fluorescence signals were photoconverted into a brown precipitate before in situ hybridization for vMHC. (I) High-power magnification of the square area outlined in H. DiI-labeled cells are brown (indicated by arrow heads in I); vMHC-positive cells stain blue; (nt) neural tube; (nc) notochord; (da) dorsal aorta; (ve) heart ventricle.
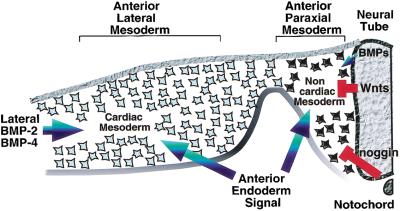
Heart formation is cued by a combination of positive and negative signals from surrounding tissues. Whereas a signal(s) from the anterior endoderm works to promote heart formation in concert with BMP signals in the anterior lateral mesoderm (blue arrows), signals from the axial tissues (red) repress heart formation in the more dorsomedial anterior paraxial mesoderm. Inhibitory signals that block heart formation in anterior paraxial mesoderm include Wnt family members expressed in dorsal neural tube (Wnt-1 and Wnt-3a) and anti-BMPs expressed in the axial tissues (i.e., noggin in the notochord). We suggest that the sum of these positive and negative signals determine the medial-lateral borders of the heart-forming region.
References
-
- Bhanot P, Brink M, Samos CH, Hsieh JC, Wang Y, Macke JP, Andrew D, Nathans J, Nusse R. A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature. 1996;382:225–230. - PubMed
-
- Christ B, Ordahl CP. Early stages of chick somite development. Anat Embryol. 1995;191:381–396. - PubMed
-
- Climent S, Sarasa M, Villar JM, Murillo-Ferrol NL. Neurogenic cells inhibit the differentiation of cardiogenic cells. Dev Biol. 1995;171:130–148. - PubMed
-
- Croissant JD, Carpenter S, Bader D. Identification and genomic cloning of CMHC1: A unique myosin heavy chain expressed exclusively in the developing chicken heart. J Biol Chem. 2000;275:1944–1951. - PubMed
-
- Frasch M. Induction of visceral and cardiac mesoderm by ectodermal Dpp in the early Drosophila embryo. Nature. 1995;374:464–467. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources